WHY THE MIND IS IN THE HEAD1 [97/98]
W.S. McCulloch
As the industrial revolution concludes in bigger and better bombs, an intellectual revolution opens with bigger and better robots. The former revolution replaced muscles by engines and was limited by the law of the conservation of energy, or of mass-energy. The new revolution threatens us, the thinkers, with technological unemployment, for it will replace brains with machines limited by the law that entropy never decreases. These machines, whose evolution competition will compel us to foster, raise the appropriate practical question: “Why is the mind in the head?”
Coming as I do between psyche anatomized and psyche synthesized, I must so define my terms that I can bridge the traditional gulf between mind and body and the technical gap between things begotten and things made.
By the term “mind,” I mean ideas and purposes. By the term “body,” I mean stuff and process. Stuff and process are familiar to every physicist as mass and energy in space and time, but ideas and purposes he keeps only in the realm of discourse and will not postulate them of the phenomena he observes. In this I agree with him. But what he observes is some sort of order or invariance in the flux of events. Every object he detects in the world is some sort of regularity. The existence of these objects is the first law of science. To detect regularities in the relations of objects and so construct theoretical physics requires the disciplines of logic and mathematics. In these fundamentally tautological endeavors we invent surprising regularities, complicated transformations which conserve whatever truth may lie in the propositions they transform. This is invariance, many steps removed from simple sensation but not essentially different. It is these regularities, or invariants, which I call ideas, whether they are theorems of great abstraction or qualities simply sensed. The reason for excluding them from physics is that they must not be supposed to be either stuff or process in the causal sequences of any part of the world. They are neither material nor efficient. So, to my mind Newton, Planck, and Jeans sin by introducing God as a sort of mind at large in the world to account for physical effects, like the action of gravity at a distance.
But let us now compel our physicist to account for himself as a part of the physical world. In all fairness, he must stick to his own rules and show in terms of mass, energy, space, and time how it comes about that he creates theoretical physics. He must then become a neurophysiologist (that is what happened to me), but in so doing he will be compelled to answer whether theoretical physics is something which he can discuss in terms of neurophysiology (and that is what happened to me). To answer “no” is to remain a physicist undefiled. To answer “yes” is to become a metaphysician—or so I am told.
But is that just? The physicist believes entropy to be somehow in or of physical systems. It may or must increase with time. But it is neither material nor efficient, in fact it is a number, namely, the logarithm of the probability of the state. It is, therefore, a measure of the disorder of an ensemble—or collection of systems. Now Norbert Wiener has proposed that information is orderliness and suggests that we measure it by negative entropy, the logarithm of the reciprocal of the probability of the state. Let us, for this argument, accept his suggestion. Ideas are then to be construed as information. Sensation becomes entropic coupling between us and the physical world, and our interchange of ideas, entropic coupling among ourselves. Our knowledge of the world, our conversation—yes, even our inventive thought —are then limited by the law that information may not increase on going through brains, or computing machines.
The attempt to quantify the information leads to a search for an appropriate unit, which, in turn, forces us to distinguish between two types of devices. In so-called logical, or digital, contrivances, a number to be represented is replaced by a number of things—as we may tally grain in a bam by dropping a pebble in a jug for each sheaf. The abacus is such a device. The nervous system is par excellence a logical machine. In so-called analogical contrivances a quantity of something, say a voltage or a distance, is replaced by a number of whatnots or, conversely, the quantity replaces the number. Sense organs and effectors are analogical. For example, the eye and the ear report the continuous variable of intensity by discrete impulses the logarithm of whose frequency approximates the intensity. But this process is carried to extremes in our appreciation of the world in pairs of opposites. As Alcmaeon, the first of experimental neurophysiologists, so well observed, “the majority of things human are two”—white-black, sweet-bitter, good-bad, great-small. Our sense organs, detecting regularities the same in all respects save one, create dichotomies and decide between opposites. These the “brain somehow fits together.” From this sprang associationalism, culminating in Mill's evolutionary hypothesis that things are similar for us which have occurred together in the experience of our progenitors, and Kapper's law that nervous structures associated in action become associated in position. Neither proposes any mechanism other than random variation and the survival of those in whom the happy concatenation occurred. We inherit a nervous system so structured that we do perceive similarities (or have ideas) and these, not isolated, but conjoined within the system in many useful ways. That synthetic a priori is the theme of all our physiological psychology, learning excepted.
How, in all these processes, can we “quantify” the amount of information? In analogical devices it is best done by examining the numerical component. They all suffer from a peculiar limitation of accuracy which can usually not be pushed beyond one part in a thousand and almost never beyond one part in a million, even in such a simple matter as weighing. Moreover, analogical devices can not be combined in any way to push the decimal point. By measuring carefully the diameter and circumference of a circle, we might analogically estimate the ratio pi to six significant figures. With a digital device, say an abacus, we can compute it to any number of places.
What characterizes a digital, or logical, device is that its possible states are separated sharply. In the simplest case there are only two. Wiener proposes that, for these bivalent systems, we define the unit of information as the decision which state it shall occupy. Notice, now, that this system has one degree of freedom—say, go or no-go—until it receives one unit of information, whereupon it has none. Next consider an ensemble of two such systems. It can be in any one of four states, and two units of information are required to match its degrees of freedom. If it were composed of three systems it might be in any one of eight states, and would require three units of information to fix it. Thus the number of possible states is 2 raised to the power which is the number of systems and each unit of information subtracts one from that exponent. Here Wiener's unit of information is exactly the logarithm to the base 2 of the reciprocal of the probability of the state, which, of course, is the negative entropy of an ensemble of bivalent systems. Neurons are bivalent systems.
Let me define “corruption” as the ratio of information in the input to that in the output. Each eye has something like a hundred million photoreceptors, each of which in a given millisecond can emit one or no impulse. In other words, it is an ensemble which can be in any one of 2100•000•000 possible states, or the amount of information it has is a hundred million units per millisecond.
Now Pitts and I have computed the information in the output of a piano player surpassing any ever known. We have given him a keyboard of a hundred keys, let him strike independently with each finger with any one of ten strengths ten times per second, and let each hand span ten keys. That sounds like a lot of information, but on computing it we find it is only about two units per millisecond.
Recent telephonic devices have sampled waves every thousandth of a second and passed on one pip if the wave was then of a given deviation from the mean, otherwise no pip. These are relayed to a smearing device and heard. It is better than 90 per cent as intelligible as the original voice. Three such pips per millisecond determined by eight possible values of the wave reproduce an orchestra. So much information at most may we hope to convey.
Whether we figure the ratio of input to output from our impossible player or from human speech, the corruption is of the order of a hundred million to one. Part of this corruption is referable to the coupling of our nervous system to our muscles and is avoided in some of the Crustacea. They use axons of several sizes, and by varying the frequency of discharge obtain more degrees of contraction than there are possible synchronous states of the nerves to their muscles. The viscosity of muscle smears the result in time, so that the rate at which impulses can come over the nerve is wasted by the inability of muscle to follow. In us a nerve of a thousand axons can be in 21000 possible states, whereas the muscle, because it can only add tensions, has only a thousand possible states. 1000 is about 210; so the corruption in passing from nerve to brawn is 100 to 1.
What becomes of all the rest of the information? To answer that, conceive neurons as telegraphic relays. Each one may be tripped by some combination of signals provided these are very nearly synchronous. It detects the coincidence and only then emits a signal to subsequent relays. Now the threshold of the photoreceptors of the eye is always varying. At any one millisecond it may be tripped by a single photon, and, at another, fail to fire in response to many. By connecting many of these to a coincidence detector set to require a reasonable number of impulses simultaneously, we have a signal which corresponds to a statistically significant fraction of its receptors and so we wash out the random variation of threshold. Thus using the relayed information that fails to agree with other information, we achieve a high probability that what goes on through the nervous system does correspond to something in the world. Perhaps it will be clearer to say it this way. The logical probability that a neuron will have an impulse in one millisecond is 1/2, that two neurons of an ensemble in the same millisecond 1/2 × 1/2. The chance that both will fire by chance simultaneously is the product of their probabilities separately; that is, it is smaller; 1/4. Therefore, in the nervous system, by repeatedly demanding coincidence we vastly increase the probability that what is in the output corresponds to something in the input. We pay for certainty with information. The eye relays to the brain about the hundredth part of the information it receives. The chance that what it does relay is due to chance is fantastically small, 2-100, a billionth of a billionth of a billionth of a tenth of one per cent.
Here, then, is the first technically important difference between us and robots. In them we cannot afford to carry out any computations, no matter how simple, in a hundred parallel paths and demand coincidence. Consequently, no computing machine is as likely to go right under conditions as various as those we undergo.
Accordingly to increase certainty every hypothesis should be of minimum logical, or a priori, probability so that, if it be confirmed in experiment, then it shall be because the world is so constructed. Unfortunately for those who quest absolute certainty, a hypothesis of zero logical probability is a contradiction and hence can never be confirmed. Its neurological equivalent would be a neuron that required infinite coincidence to trip it. This, in a finite world, is the same as though it had no afferents. It never fires.
In all of this I take it for granted that you are familiar with the all-or-none law of the nervous impulse, with the brevity of latent addition, with the duration of synaptic delay, with the evidence for spatial summation and for inhibition at a synapse, and with the local origin of energy from metabolism, all of which together insure that the principal circuit actions of the nervous system are those of the digital, or logical, kind. My reason for letting time flow in lapses of a millisecond is based on the work of Lorente de Nó, who has given us some of the best measures of the other properties of conduction. It is probable that no neuron can sustain more impulses than a thousand per second, even under the healthiest conditions, and one millisecond will include synaptic delay, or absolute refractory period, or the front of the pulse itself. These permit us to treat the nervous impulse as an atomic event.
But a nervous impulse is also a signal. It is true if what it proposes is true, otherwise it is false. It is false if it arises from any cause other than the adequate, or proper, excitation of the cell. The threshold of the dark-adapted eye for light is about a photon in several seconds. Pressure applied to the eye will evoke impulses, but the energy required is many million times more. Press on the eye and you see light when there is no light. The signals are false. Thus nervous impulses are atomic signals, or atomic propositions on the move. To them the calculus of propositions applies provided each is subscripted for the time of its occurrence and implication given a domain only in the past. In terms of such a calculus applied to nervous nets, Pitts and I have been able to prove that even nets devoid of circles can realize any proposition which is a logical consequence of its input. As this is the most that any net can do it is obviously an adequate theory. We know, of course, that facilitation and extinction occur, and we showed that whatever these can effect can be done digitally, or discretely, by go, no-go devices. In our first essay, we were unable to obtain much more than the calculus of atomic propositions; but, by introducing circles in which a train of impulses patterned after some fact could circulate, we did get existential operators for time past.
This is the argument: In a net in which there are no re-entrant paths a signal anywhere in the net implies a signal in a neuron nearer to receptors, and so backward in time until we arrive at the receptors. The signal here and now in this net implies the signal sent there just then. But once set going, a disturbance in a closed chain implies that there was a signal in its input at some time but does not indicate at what time. In short, the reverberating activity patterned after something that happened retains the form of the happening but loses track of when it happened. Thus it shows that there was some time at which such and such occurred. The “such and such” is the idea wrenched out of time.
It is an eternal idea in a transitory memory wherein the form exists only so long as the reverberation endures. When that ceases, the form is no longer anywhere. Only this kind of memory remains to aged brains in which no new abiding traces can be made and old ones fade. While we are young, use leaves some sort of change, as freshets cut their channels in the hills so that aftercoming waters follow and enlarge their beds. Yet all other forms of memory, including written records, do nothing which cannot also be achieved by mere reverberation, and hence add nothing to the theory.
There are other closed paths important in the origin of ideas, circuits which have “negative feedback.” In terms of them reflexes were first defined as actions starting in some part of the body, setting up impulses to the central nervous system, whence they were reflected to those structures in which they arose, and there stopped or reversed the process that gave rise to them. All inverse feedbacks have this in common, that each establishes some particular state of the system, for they bring it back toward that state by an amount which increases with their deviation from that state. They are, as we say, error-operated. The state toward which they return the system is the goal, or aim, or end in and of the operation. This is what is meant by function. On these circuits Cannon founded his theory of homeostasis, and Rosenblueth and Wiener their theory of teleological mechanisms.
Any such circuit becomes a servomechanism as soon as the particular state it is to seek can be determined for it. Thus the stretch reflex tends to keep our muscles at constant length, but that length is determined for these circuits by more complicated arcs which traverse almost all parts of the central nervous system and require the reflex to seek those states which permit us to stand and move.
One reflex turns the eyes toward anything that enters the visual field. Its path runs from the eye by fibers that bypass the geniculate to enter the superior colliculi upon which they map the visual field. Here local circuits compute the vector from the center of gaze to the center of gravity of the apparition and send this information to the oculomotor nuclei which, in turn, relay orders to the appropriate muscles and turn the eyes so as to decrease that vector. As it reaches zero the eyes come to rest with the apparition centered. This reflex, I am told, will operate even in a man who has lost one-half his visual cortex, if he is dark adapted and a light, unseen by him, is placed in his blind field. If two are placed there the eyes turn toward a position intermediate. Under these conditions but with cortex intact, the eyes turn similarly but then snap from spot to spot, for the reflex is then subservient to impulses from the cerebrum. By turning the eyes so as to center the form, the reflex rids the apparition of the gratuitous particularity of the place at which it appeared. Every reflex, by running through a series of intermediate states to that established by it, rids some item to be observed of some fortuitous specificity. In the case of the collicular reflex, it has selected the centered form from among all possible exemplifications. Once in this, the canonical position, the system is ready for the computation of the form. There is little doubt that in us this computation occurs in the cerebral cortex, notably the visual areas. What happens then as the eyes turn rapidly is a series of on and off signals from most portions of the eye. These serve to clean the slate for the centered form unencumbered by blurring due to motion. The latter we suffer only when the eyes turn slowly; for instance, when we are very tired.
There are negative feedbacks within the brain. One of these resembles the automatic volume control in the radio. It tends to keep constant the sensory input to the cerebral cortex. In so doing it gives us another existential operator, for it detects that there was some intensity such that it was of this-or-that figure. In the case of vision, this circuit follows two other devices serving the same end; namely, the slow adaptation of the retina and the rapid change of pupillary diameter. All together these enable us to detect the form though the intensity of its illumination range through 39 decibels, that is, from faint starlight to full daylight—only we may not look at the sun without closing our eyes.
There are also appetitive circuits with a part of their path, from receptors to effectors, inside us, and the rest through the world outside. They are said to be inverse feedbacks over the target. Given any two inverse feedbacks which working together would destroy us, like swallowing and inhaling, there is built into us some connection between their paths whereby, when both are set going, one stops the other. In the case of learned modes of appetitive behavior, similar inhibitory links must be acquired or we perish. If we have three incompatible circuits in which A dominates B and B dominates C, the chances are equal whether A will dominate C or C, A. We speak of the end-in-operation of the dominant as of greater value. We have even tried to construct scales of value for diverse ends, but, since dominance is sometimes circular, values are not magnitudes of a single kind, and the terms “greater” and “less” are simply inapplicable. What we have called the value anomaly and regarded as evidence of a lack of order or system bespeaks, in fact, order of a kind we had not imagined, and a system tighter knit. Here endeth the psychological blind alley and Plato's theory of the Good. We cannot make one scale of value that predicts choice. Only knowledge of mechanism permits such prophecy.
I drew a circuit to move a figure, given anywhere in a mosaic of relays, to all positions in one direction. From each relay impulses ascended diagonally in the required direction through sheets of relays resembling the original mosaic and so spaced that their constituent relays formed columns perpendicular to the planes. And I set the threshold of all relays so that none would fire except when a slanting impulse coincided with one in the plane of that relay. I brought the output of every relay vertically all the way down to the original mosaic. Now when there is a simultaneous volley of the required figure at the given place in the original mosaic and at the same time a simultaneous volley to all the relays of any one of the sheets above, the figure is reproduced on that sheet by a volley in the relays where the slanting volley hits the sheet. Thence it projects straight down on the original mosaic. This reproduces the figure at a distance which steps off in the direction of the slant by a number of relays proportional to the height of the excited sheet. Now let the figure endure by a series of volleys at its origin, and excite the sheets successively upward, and the figure will be translated step by step from the origin to all possible positions in the direction of the slant. Whatever shape is present in the input to this circuit is preserved in these successive representations and, as the output descends vertically, the shape is translated without distortion. Von Bonin, who had worked with me on the auditory cortex, when he saw the diagram mistook it for a drawing of that cortex. Certainly we had but to replace the relays by pictures of neurons and the similarity was startling. The parallel functions are even more alike. We can center a form seen by turning our eyes, but there is no way we can tune our ears so as to translate a chord up and down the scale. Our brain receives it at a fixed key or pitch. In the primate these pitches map longitudinally on the input to Heschl's gyrus, so spaced that octaves span nearly equal distances. If this, the primary auditory cortex, worked like my circuit it would move the output up and down the axis of pitch while it preserved the interval, and so the chord. Here is an existential operator for chord regardless of pitch. The output asserts that there were pitches such that there was this or that chord.
Is there anything in physiology corresponding to the sequential excitation of the sheets? And, if so, how fast can it complete a cycle? There is the familiar alpha rhythm of the cortex, a shift of voltage that rises and falls through the cortex ten times per second. Although the correspondence may be entirely fortuitous, this is about the rate at which chords can be distinguished—ten per second. Now we need excellent histological studies by the Golgi method to know whether the detailed connections of cells in this area are what the hypothesis requires. These must be made in specified planes to match our physiological data. Because incoming signals and outgoing signals, like the pulse of scansion, ascend and descend through the cortex, when the cortex is at work the sweep of scansion should disappear, as it does, in the twinkle of details.
There are at least two ways that the output of this primary cortex may convey a chord regardless of pitch. There may be an inverse feedback which stops the figure of excitation when it reaches a canonical position along the axis of pitch, but there is no evidence that this exists. The other way is suggested by anatomy. Beneath the receptive layers of the cortex are columns of cells where properly timed impulses may be accumulated through a time equal to the sum of their synaptic delays to coincide upon efferent cells whose axons go to the adjacent, or secondary, auditory cortex.
If they terminate there at random, and if the cells there merely require coincidence to fire, we will have for every chord regardless of pitch a corresponding spot of maximal coincidence. The activity of this spot proposes the required universal, or idea. If we were to excite this spot electrically in waking man at operation, he should report hearing the chord. He does, but unfortunately no one has asked him whether he hears it at some particular pitch. The experiment is difficult because the primary auditory cortex is buried deep, and the secondary adjacent to it almost as deep, in the fissure of Sylvius. Moreover, I have not been able to map well the projection of the primary upon the secondary; and, finally, the interpretation is complicated by a direct projection upon the second with the sequence of pitches reversed. Fortunately these difficulties are not present in the visual area of man or monkey.
I drew a circuit to extract shape regardless of size, and this was mistaken by both von Bonin and Percival Bailey for a schematic representation of the outer strip of Baillarger which makes the visual cortex the “area striata.” We start again with a mosaic. Select a point to represent the center of gaze and map the visual field as a set of concentric circles whose radii are proportional to the logarithm of the angles at the eye. From the mosaic let impulses proceed along branching channels spraying outward as they ascend through sheets of relays in which the density of relays decreases but their threshold increases as we go from below upward. From all of these relays let signals rise to corresponding upper layers of relays where coincidence with sweeping pulses is required, and let the signals of these layers converge on relays of low threshold, thence descend to leave the area striata. Now, with the pulse of scansion we shall have successively in this output all possible dilatations and constrictions of any figure in the input. The possibilities are limited by the grain and gross dimensions of the cortex, but these limit input and output equally. Since we have, in the output, all sizes made from a given one it makes no difference which size was given in the input.
Had we not conformed to present knowledge by mapping radial angles by their logarithms we would be compelled to require that the branching ascent of the input take a radial direction, but as it is it may branch nearly equally in all directions. Hence we cannot hope to detect much difference in histological study, even by silver stains of fibers, between sections cut radially and others, tangentially, in the visual cortex, or even in ones parallel to the surface. Thanks to Ramon y Cajal and Lorente de Nó, we know that the anatomical connections are at least sufficient for the theory. Here, as in audition, if the alpha rhythm evinces the scansion, we should be able to see ten forms per second. We can. Faster, they blur, merge, or glide into one another. Moreover, a rise of metabolic rate with fever or hyperthyroidism causes a rise in alpha frequency and a rise in the number of distinguishable frames per second.
When strychnine is applied locally to a spot on the cortex it causes the cells there to fire almost in unison. The fibers leaving that area then carry nearly synchronous volleys of impulses. These can be traced to the ends of the axons if there are enough of them near together. When these axons turn up again into the cortex anywhere, we can detect them there as a sharp change in voltage, the so-called “strychnine spike.” When Dusser de Barenne and I strychninized a pinhead spot on the area striata, strychnine spikes appeared at many points in the secondary visual area as if the output from each spot in the area striata were scattered at random in the secondary visual area. Hence, from any particular set of spots in the primary area there will arise by chance some spot of maximum excitation in the secondary area. Activity at this spot implies activity in some figure of spots output by the primary; hence some shape regardless of size. Electrical stimulation of a spot on the, primary visual cortex in waking man is reported by him as a blurred circle of light, whereas similar excitation of a spot in the secondary area is reported as a form. Moreover, this form, while it has a position in space, in the sense that he can point at it, has none in the visual field. Nor does it seem to have size there, any more than the recalled image of the moon seems to subtend one particular angle at the eye.
The mechanism we propose for abstracting chord and form is really computing a kind of average, and that average will not be seriously affected by small perturbations of excitation, of threshold, or even of particular connections as long as they are to cells in the right neighborhood. This conforms to clinical findings. A man may have several holes in his visual cortex, as big as or bigger than, the hole in his retina called the optic disk, and, except in a small number of cases, the forms seen will be unaffected. Although in such a case we can map these blind spots, he will not see them and the things seen will appear to be continuous through the blind spots. Scrutiny of the hypothesis even suggests that this process may account for much relational determination, for the four corners of a square in the input would be completed as a square, whereas parts of the sides might well flop, seeming now a Maltese cross and now a square, etc. These flops would be the outcome of rivalry between two maxima for dominance over subsequent areas. Thus, for vision, our hypothesis fits well all known facts.
Older schools of physiological psychology and of neurology, guided by atomistic associational doctrines, tended to think in terms of neurons, each of which had one duty, for example, to know squares. This seems to be at least partially true of spots in the secondary visual cortex. Gestalt psychologists have treated the mosaic of relays of the cortex as if it were a field on which sensations mapped synchronously. This seems more likely true of receptors like the retina, for even its cortical replica is bisected by a line down the middle of the field and the halves mapped far asunder. Now it is easy to show that both of these “caricature” the nervous process. We need only note that a nervous net can take any figure in space, requiring an ensemble of a given number of neurons simultaneously, and convert it into a figure of impulses over a single neuron requiring as many relay times as there were neurons in the ensemble, and vice versa. From this alone it is clear that we cannot tell what kind of thing we must look for in a brain when it has an idea, except that it must be invariant under all those conditions in which that brain is having that idea. So far we have considered particular hypotheses of cortical function. They are almost certainly wrong at some point. Because they have already had to fit many disparate data, they are of little a priori probability. They prophesy the outcome of an infinite number of experiments, some of which are almost certain to refute them.
But with respect to the underlying theory, which is merely glorified tautology, there is no such possibility. It is, in fact, little more than a simple application of the theory of groups of transformations. For any figure in the input of a computing machine it is always possible to calculate an output invariant under a group of transformations. We calculate a set of averages, for all members of the group, of numerical values assigned by an arbitrary functional to each transform of the information conceived as the distribution of excitation at all points and times in an appropriate manifold. To define the figure completely under these transformations, we would need a whole manifold of such averages for various functionals, and this manifold would have to have as many dimensions as our original one; but, for practical purposes, we usually need only a few averages. Since in the finite net of relays the number of transformations in finite time is finite, we may use simple sums instead of averages.
This general theory describes all processes of securing invariants, or having ideas, which we have discovered or invented to date; and one mechanism differs from another in the nature of its arbitrary functional. For example, in the cerebral circuits proposed the functional may always assign the value one to any vector in the manifold if the particular point had a signal in the previous relay time, and, if not, assign it zero, whereas, in the reflex circuit for centering an apparition the functional clearly depends upon the figure of excitation in the manifold, and changes as the form centers; in effect, it assigns the value zero to all save the last transform on the cortex.
We may, of course, make the output of any calculator of invariants (or of several of them) the input to another and so have an idea of ideas, which is what Spinoza calls consciousness, and thus get far away from sensation. But our most remote abstractions are all ultimately reducible to primitive atomic propositions and the calculus of the lowest level. The domain of their implication lies only in time past. If their domain extended into the future, our sensations would imply our thoughts and our thoughts imply deeds. They do not, for even if the threshold of every cell in the nervous system were fixed, between the time we conceive an act and the time the impulses reach the motor horn cell, other signals from the world may get there first, and so often thwart us. We note the failure in the fact and are forced to distinguish between what we will and what we shall do. Hence the notion of the will.
But we do guess at things to come. When we run to catch a baseball we run not toward it but toward the place where it will be when we get there to grab it. This requires prediction. We behave as if there were some law compelling the world to act hereafter as it did of yore. Only one of our predictive circuits has been carefully studied by physiologists. It is responsible for optokinetic nystagmus. It has a tendency to persist, which may be seen when a train stops, for it then attributes motion in the opposite direction to the ties and rails. The earmark of every predictive circuit is that if it has operated long uniformly it will persist in activity, or overshoot; otherwise it could not project regularities from the known past upon the unknown future. This is what, as a scientist, I dread most, for as our memories become stored, we become creatures of our yesterdays—mere has beens in a changing world. This leaves no room for learning.
Neurons are cheap and plentiful. If it cost a million dollars to beget a man, one neuron would not cost a mill. They operate with comparatively little energy. The heat generated raises the blood in passage about half a degree, and the flow is half a liter per minute, only a quarter of a kilogram calorie per minute for 1010, that is, 10 billion neurons. Von Neumann would be happy to have their like for the same cost in his robots. His vacuum tubes can work a thousand times as fast as neurons, so he could match a human brain with 10 million tubes; but it would take Niagara Falls to supply the current and the Niagara River to carry away the heat. So he is limited to about the thousandth part of man's computer. He has to be very careful to specify in detail which relays are to be connected to a given relay to trip it. That is not the case in human brains. Wiener has calculated that the maximum amount of information our chromosomes can convey would fill one volume of the Encyclopedia Britannica, which could specify all the connections of ten thousand neurons if that was all it had to do. As we have 1010 neurons, we can inherit only the general scheme of the structure of our brains. The rest must be left to chance. Chance includes experience which engenders learning. Ramon y Cajal suggested that learning was the growing of new connections.
I do not doubt that the cerebral cortex may be the most important place in primates. But it is certainly the most difficult place to look for change with use. Think of it as a laminated felt of fibers which serve to associate neighboring rough columns of cells nearly a hundred high and linked together vertically by their axons. These columns are then connected to distant columns by axons which dip into the white matter and emerge elsewhere into the cortex. These last connections I have studied for many years but have at best a general picture of how areas are related, certainly nothing that could give the detail necessary to distinguish between its connections before and after learning.
To understand its proper function we need to know what it computes. Its output is some function of its input. As yet we do not know, even for the simplest structure, what that function is. We have only a few input-output curves for the monosynaptic reflex arc obtained by David Lloyd, and now a few more by Arturo Rosenblueth. Walter Pitts is analyzing them mathematically at the present moment and has as yet no very simple answer. There is no chance that we can do even this for the entire cortex. That is why we need such a hypothesis, as we have proposed for particular areas, for these may be disproved by records of electrical activity recorded concurrently at a few specified places.
Contrast our ignorance of its proper function with the detailed present knowledge of the projection of the sensory system upon it. For on at least two-thirds of its surface we can map the surface of the body, outside, and, to some extent, inside, so as to assign to every square millimeter of cortex the origin of its specific afferents and through them the exact position of the organs of sense. Beginning last summer, and continuing right now, the surface of the cerebellum, upon which the body maps similarly, is being stimulated and its projection to the so-called motor and sensory cortex, primary and secondary, explored and plotted millimeter by millimeter. Also now the projections of so-called non-specific afferents are receiving similar attention. Thus within a year or so we will know the geometry of its input and will be ready to seek in loci well defined the temporal pattern of its input.
I wish we could say half as much for our knowledge of its output. Since the days of Bubnoff and Heidenhain it has been electrically stimulated and the resultant change in muscle and gland carefully observed and elaborately recorded. But these responses depend upon the state of all subservient circuits which have yet to be analyzed. Hardly a month passes but what we are confronted by surprises. Frequency as well as shape of electrical pulses have been shown to determine the very path of the descending pulses from one and the same cortical focus. For example, volleys of impulses from the so-called face motor cortex, if more than ten or twelve per second, play principally through the nucleus of the seventh nerve upon the muscles of the face; whereas, if less than ten per second they are relayed almost exclusively through the nucleus of the twelfth to the tongue. Finally, the response to one and the same form of stimulation of a single focus in the motor cortex for one limb is determined both in amplitude and in direction by the motion and by the position of the limb at the time of stimulation. Clearly, to understand the significance of the output of the cerebral cortex we must know, for every subservient structure, the input-output curves. Even that will not be enough, for when several of them form a re-entrant circuit we must know their relations. Until we do so we will be in danger of attributing to the cerebral cortex functions proper to lower structures.
Last, but not least, the cortex is itself part of many re-entrant systems, and what our hypothesis attributes to cortex alone in securing invariants, or having ideas, may well depend upon loops joining it and the thalamus. From all these uncertainties I would turn to something simple as the monosynaptic arc of the stretch reflex and, by procedures far from normal, try to teach it something. It will be difficult, for in it the connections are as certainly determined as in a man-made computing machine; and we will have to break old connections before we can form new ones.
This brings us back to what I believe is the answer to the question: Why is the mind in the head? Because there, and only there, are hosts of possible connections to be formed as time and circumstance demand. Each new connection serves to set the stage for others yet to come and better fitted to adapt us to the world, for through the cortex pass the greatest inverse feedbacks whose function is the purposive life of the human intellect. The joy of creating ideals, new and eternal, in and of a world, old and temporal, robots have it not. For this my Mother bore me.
References
Barker, S. H., and Gellhom, E. Influence of suppressor areas on afferent impulses. J. Neurophysiol., 1947, 10, 125-132.
Bell, Charles. On the nervous circle which connects the voluntary muscles with the brain. Proc. Roy. Soc., 1826, 2, 266-267.
Cajal, Ramon y, S. Histologie du systeme nerveux. 2 vols. Paris: Maloine, 1909, 1911.
Lorente de Nó, R. Sections in J. F. Fulton's Physiology of the nervous system. London: Oxford University Press, 1943.
Maxwell, C. On governors. Proc. Roy. Soc., 1868, 16, 270-283.
McColl, H. Servo-mechanisms. New York: D. Van Nostrand Co., 1945.
McCulloch, W. S. A heterarchy of values determined by the topology of nerve nets. Bull. of Math. Biophys., 1945, 7, 89-93.
McCulloch, W. S. Finality and form. Fifteenth James Arthus Lecture, New York Academy of Science, May 2, 1946.
McCulloch, W. S. Machines that think and want. Lecture at the American Psychological Association, September 9, 1947.
McCulloch, W. S. Through the den of the metaphysician. Lecture at the University of Virginia, March 23, 1948.
McCulloch, W. S. Teleological mechanisms. Ann. New York Acad. Sci., 1948, 50, 4.
McCulloch, W. S., and Lettvin, J. Y. Somatic functions of the central nervous system. Ann. Rev. Physiol., 1948, 10, 117-132.
McCulloch, W. S., and Pitts, W. How we know universals. Bull. Math. Biophys., 1947, 9, 127-147.
McCulloch, W. S., and Pitts, W. The statistical organization of nervous activity. J. Amer. Statistical Assoc., 1948, 4, 91-99.
Rosenblueth, A., Wiener, N., and Bigelow, J. Behavior, purpose, and teleology. Philos, of Science, 1943, 10, 18-24.
Wiener, N. Cybernetics. New York: John Wiley and Sons, 1948.
Wittgenstein, L. Tractus Logico-Philosophicus. London: Paul, 1922.
Discussion
DR. LORENTE DE NÓ: The main question in our minds is whether the theory as a whole is going to stand or not. I think that probably many of the details will not stand, but that the main concept will certainly remain. I'm quite sure that all of my colleagues will agree that Dr. McCulloch has brought what we know of both the anatomy and the physiology of the brain closer to an integrated whole than it has ever been before, and I want to congratulate Dr. McCulloch very much and very sincerely.
DR. VON NEUMANN: I would not like to attempt a detailed discussion of the very beautiful and very interesting presentation made by Dr. McCulloch, perhaps something like that can be done in the general discussion. I will, however, ask two questions, both dealing with only one aspect of the matter. You have emphasized that you are giving sufficient mechanisms and that it is in conflict with your entire philosophy at this time to claim that these are necessarily the ones that are used. You give proofs of possibility. There is, nevertheless, one point where the question of the actual mechanism is especially burning, and that is the question of memory. You have pointed out that there are positive feedbacks—reverberating circuits—built out of switching organs which are quite adequate as memory. If there were nothing else in the world except neurons, you could build memory out of neurons. My own feeling is that if one were really to construct in this way a nervous system with its known attributes it would probably take more neurons than there are, but this is an aside. My real question is this: First of all, I have observed that all neurologists seem very certain that the reverberating circuit trick is not used in making the actual memory. Amorphous intuition points in the same direction. In surmising this is not so, I have always had a bad conscience. I am not sure why they are so positive. What is the best evidence one can give for this?
The second question is this. Most neurologists with whom I have had an opportunity to talk seem to be equally convinced that memory is due to some lasting changes somewhere on the body of the nerve cell, somehow connected with alterations of thresholds. Is it not better to say that there probably is a memory organ somewhere, but that we are absolutely ignorant as to where it is—probably as ignorant as the Greeks, who located the whole intelligence in the diaphragm?
DR. McCULLOCH: I'm afraid my answer is necessarily a bit lengthy. In the first place, I would like to contrast as sharply as possible the maximum length of what I consider reverberative memory, with the enduring memories which we bring on from childhood. I have seen a man over 80 years of age walk into a meeting of a Board of Directors and for 8 hours work out from scratch all of the details necessary for the sale of a complete railroad. He pushed the other men so as to get every piece of evidence on the table. His judgment was remarkably solid. The amount of detail involved in the transaction was enormous, and it actually took over 6 hours to get all of the requisite details on the map. He summarized that detail at the end of the meeting, in a period of a half an hour, very brilliantly, and when he came out he sat down, answered two letters that were on his desk, turned to his secretary, and said, “I have a feeling that I should have gone to a Board of Directors Meeting.” He was not then, or at any later time, able to recall one iota of that meeting, and he was in that state for nearly a year before he died. This is the picture of what we call “presbyophrenia.” In that state, whatever our memory organ is, we are unable to make any new record in it. Actually, the recent paths begin to fade, leaving only earlier memories to pop up. So, at that period, one is more likely to remember in detail and individually, the things that happened in childhood rather than the things of later years. Such a memory goes, quits, stops, the minute the brain is used for something else, or the minute that it comes to rest. Here is a span of at least 8 hours of high cerebral activity in carrying the details from the first moment of the meeting to the end of the meeting.
Per contra, not all memory can be of this reverberative kind. It is obvious that, although this kind of memory is carried reverberatively in the brain, it cannot endure during very deep sleep and it cannot endure during narcosis. It goes out, when the brain has a seizure and it goes out in sleep. In the one case, it goes out because the whole apparatus is pervaded by what I will call shock waves which go through it and through it and through it in the fit. In the other case, it goes out because it has no signals traveling—the brain is “shut down.” I believe that only lower mechanisms are really busy in deep sleep. Now then, why do we want to attribute the memory to the brain at all? Why may it not be in the spleen or somewhere else? The answer is because injuries of the brain, but not injuries of other things, do result in losses of memory, and that is the fundamental reason for pinning it on the brain.
The next question is: Why does one attribute this, the enduring memory, to a growth process—change with use—somewhere in the excitability in neurons rather than elsewhere? Well, first, because it is a relatively lively process, and when things are growing, one tries to pin it on growth processes. Second, because it has the peculiarity that what we learn later is only a modification of what is already laid down. It is an accumulative affair of this sort. Why attribute it to the junctions of cells? Because there is where we imagine the switching takes place, and this is the kind of evidence on which we base it.
Let me tell a tale out of school, even if future evidence fails to support it. I will ask Dr. Lettvin's forgiveness later. The theory and experiments are his, although I have done some of them with him. His theory, and experiments, are designed to meet the requirements of the conditioned-reflexologists, Pavlov et al. In deference to them we will name one source of afferents U, the unconditioned afferents, which can excite an efferent R, the responsive motoneurons, and a second set of afferents C, the conditioned afferents, and—Heinrich Klüver and Warren McCulloch to the contrary notwithstanding— we will forget for the moment “stimulus equivalence” or “universals secured by averaging over groups of transformation” and treat U, C, and R as individual neurons. U can always fire R; but C can become able to do so reflexologically only if C and U are excited so as to be active concurrently. I mean that if both are concurrently active then, thereafter, C alone shall be able to fire R. Now the gist of Dr. Lettvin's analysis is that it still further simplifies the required assumption. He asks, for what do we need U—except to excite R—so why not make the simple assumption that if C and R are simultaneously active, C shall become able to fire R? Naturally this simplification should not occur to a psychologist for he has to use U to excite R. But a physiologist may put his electrodes directly upon R, if he can get them there, or he may fire it antidromically.
Now it is clearly established by the surgically and electrically perfect experiments of Donald Marquis and Arthur Ward that the intact spinal cord cannot be conditioned. But Culler and Shurrager did sometimes obtain conditioning of what they believed to be the two-neuron reflex arc, and this in experiments on by no means their technically best preparations. Wiener's theory that the spinal cord has suffered a “Wärmetod” of information by the time we are able to walk (that is, its connections are all soldered in) would account for this discrepancy. One has only to suppose that one must destroy something ending on a motoneuron to leave root room for another afferent; and the cord, no longer intact, could be conditioned even as Cajal supposed, by something else making connection with the motoneuron.
Moreover, the technically impeccable experiments of David Lloyd have proved that of two muscle antagonists at a single joint, each by its afferents (from stretch receptors) inhibits, and only inhibits, its antagonist. They show further that this inhibition occurs at the synapse on the motoneuron without time for internuncial intervention.
It follows, that if one were to cut the dorsal roots of the nerves for extension at the knee, there would be root room on its motoneurons for afferents from the flexors of the knee to get a greater hold on these motoneurons. And Dr. Lettvin's ingenious theory of synaptic transmission predicts that, if this happens, instead of inhibiting the motoneuron, these afferents will then excite them.
This, in substance, is what we did. We cut the dorsal roots of the extension reflex, stimulated its ventral roots antidomically, and at the same time stimulated the flexor muscle nerve. This we did to both for a long time, minutes or even hours, at about forty per second, from separate stimulators. Thereafter, but not before, threshold stimulation of the flexor nerve elicited contractions of the extensor. Thus Dr Lettvin has proved the cord, no longer intact, can be conditioned, but not quite as the psychologist would have it – for he stimulated C and R, not C and U, concurrently. He has not as yet published, and will not publish, these findings to physiologists until his records of the times of these impulses at dorsal and ventral roots show conclusively whether or not this functionally new path is monosynaptic.
If we are not misled by the sensitivity of denervated structures, and the cord does so learn, this is crucial to psychologists. The cord is a sufficiently simple structure and is sufficiently well known for us to hope that n anatomist, with some new technique, may be able to find structural changes.
From what I have said, it should be clear that I do not think learning normally occurs in the spinal cord. Even in the earthworm learning seems to reside, albeit not in the most anterior segments, still in the forward ganglia. In mammals it may be in the midbrain, or even in the cortex, but our chances of locating anatomical changes there are negligible.
DR. VON NEUMANN: The experiment which you described – if it were done, and if the time relations were clear – would be very convincing. In this case one could at least feel certain that the “conditioning” consists in a physical change in the cell actually under consideration.
The “reverberating circuit” model for the memory does not strike me, as I said before, as a particularly elegant one. Nevertheless, it is important to know whether it is a possible model or not. I understood you to state that it is not. What is the decisive argument against it? I understood it to be that there are states when one can be fairly certain that the cortex is totally inactive and yet memory persists. What exactly is the evidence for this “total inactivity” of the cortex? Is it that one has not so far succeeded in picking up any electrical signals from it?
DR McCULLOCH: That’s right. When our amplifier is turned up maximally, we pick up activity only from the respiratory mechanism and similar structures. Only they keep on going in deep sleep and in the coma following seizures.
DR. VON NEUMANN: Is this reliable enough to know that there is nothing else there?
DR. McCULLOCH: Well, let's take the more powerful case — that in which you have a seizure with tremendous waves of signals through the works.
DR. VON NEUMANN: Does one know that they are really going through all channels?
DR. McCULLOCH: I think, pretty firmly, yes; I don't believe any part of the nervous system is unaffected.
DR. VON NEUMANN: The organism is fairly well set up to protect certain parts, is it not?
DR. McCULLOCH: The grand mal convulsion is the occasion on which that protection breaks down.
DR. VON NEUMANN: It does not seem to for memory.
DR. LORENTE DE NÓ: The difficulty in making memory reverberating paths is chiefly this: To maintain a steady state in any kind of reverberating path, the closed chains of neurons are arranged so that either you have incremental activity or decremental activity. Either the thing begins spreading to involve more and more and more neurons, or it decrements, after coming to a maximum, and then decays and disappears. Probably the secrets that Dr. McCulloch just gave us can be compared with the gramophone record. While we are playing a gramophone record, we have an articulation of impulses; later, when the record is over, memory is deposited in a different manner. As you listen to what I am saying now there are a lot of circuits operating according to those principles; but, then, when in a moment I stop talking, those signals will have stopped and memory will be somewhere else, in some other area.
DR. VON NEUMANN: I see the plausibility of what you say, but I still have a residue of uncertainty left. Your arguments about electrical circuit analogies are plausible, but they are nevertheless influenced by our particular kind of experience in this field. Your judgment based on anatomical experience is perhaps more cogent. It may be anatomically established that closed (and hence potentially reverberating) neural pathways do not exist in the necessary, vast numbers.
Another comment I would like to make is this. I see an argument that one might make against the view that memory in any form actually resides in the neurons. It is a negative argument, and far from cogent. How reasonable is it? This is the argument: There is a good deal of evidence that memory is static, unerasable, resulting from an irreversible change. (This is of course the very opposite of a “reverberating,” dynamic, erasable memory.) Isn't there some physical evidence for this? If this is correct, then no memory, once acquired, can be truly forgotten. Once a memory-storage place is occupied, it is occupied forever, the memory capacity that it represents is lost; it will never be possible to store anything else there. What appears as forgetting is then not true forgetting, but merely the removal of that particular memory-storage region from a condition of rapid and easy availability to one of lower availability. It is not like the destruction of a system of files, but rather like the removal of a filing cabinet into the cellar. Indeed, this process in many cases seems to be reversible. Various situations may bring the “filing cabinet” up from the “cellar” and make it rapidly and easily available again. There are many examples of this: the “forgetting” and subsequent “remembering” or recovering of languages, telephone numbers, names—paralleling the decreased or increased need for their use.
This organizational situation is a very plausible one, if there is a memory which is much larger than the available switching facilities for its selective use. Indeed, if the memory is thus larger than its switching system, it will be necessary to introduce a system of priorities for various parts of the memory. Each part may then, upon occasion, be moved into regions with rapid accessibility, or into regions with less rapid accessibility. Or, rather, it may not be moved from region to region, but be connected to quickly or to less quickly functioning portions of the available switching system.
If this is so, then the memory cannot reside in the actual switching organs in the neurons, and its capacity must be much greater than that represented by the switching system. One must then postulate a very high-capacity memory organ or organization, with considerable bottlenecks at the “input” and “output,” that is, at the points of contact represented by the switching system.
Does this sound plausible, or is there some flaw in my argument?
DR. BROSIN: May I break here with the tradition of immediate reply? This is a very large subject, and I would like to see if there are other commentators. You may gather the evidence, and if we do not have time enough today, you can have a full dress performance tomorrow.
DR. GERARD: I would like to ask a few questions, some of which have already been touched upon. I have a very trivial one first. I didn't quite see why you place such disparate emphasis on the manipulation of the output of the brain in efferent systems and paths, as compared to the problem of the manipulation of the input of the brain. If I correctly understood you, you are not particularly worried about the input side. I don't see why that is, and I would like to have you explain it a little more clearly.
I have, also, two other points that touch on this memory problem. If learning and remembering are based on growth processes of some sort, then they should not be basically different from developmental and maturational behavior; and yet it seems to me that some of the most striking experimental work in the past does emphasize a very fundamental difference between the maturational learning in the nervous system and acquiring a new behavioral capacity—experiential acquiring of new behavior possibilities. The former takes place certainly without any external experience, but you can see that there is internal experience. On the question of rest and activity of the nervous system, several members of the audience, during our intermission, raised the question with me whether you are not neglecting something that you might call automatic activity of neurons. The assumption is that the output will be determined by the input, rather than by something happening independently of the input. I will put the question to you in this way. Do I correctly assume that you were suggesting that the scansion machine in your projection area mechanism is the spontaneous brain wave, and whether it starts there or below is immaterial? I would like to have that elaborated and made a little bit clearer. I personally am surprised at the answer you gave Dr. von Neumann, that the brain can be completely quiet. I don't believe that electrically, or in any other way, it is ever completely quiet at any time except in death.
DR. MC CULLOCH: I said, except for lower mechanisms.
DR. GERARD: I believe that, even in the other mechanisms, I have never seen a completely silent brain.
DR. McCULLOCH: No, I don't believe that brain matter is ever completely quiet. I'll take care of that question later. For the moment it is enough that there are times when no signals are reverberating.
DR. GERARD: If there is a separate memory organ, along the line of Dr. von Neumann's comment, in which you have your files easily accessible or down in the basement, that would argue against the memory traces being associated with the neurons themselves throughout the brain. What about the reversible amnesia problem, where all past memory vanishes for long periods and then comes back again? If learning involves the establishment of new functionally effective connections between neurons in the brain (whether by growth, by physiological change in threshold, or what not), and if that depends on activation of neurons and association with experience, then it seems to me that it should follow that if the threshold of neurons is held low, just in general during the experiencing of experience, learning should be enhanced. Dr. Lettvin raised the level of excitability of neurons in the nervous system. There is more chance of a particular input leaving a permanent modification, or even a temporary one. I hope some of the psychologists here can bring the evidence in, but I don't know. However, as far as I'm aware, conditioning under the influence of stimulating drugs has not changed the rate of this conditioning. '
The last question I should really leave for Dr. Lashley, since it is in his field. If these networks of neurons (even allowing for considerable interchangeability of particular elements of the net) are organized so beautifully in the striate and elsewhere for these particular functions, then how do you account for some of Dr. Lashley's critical experiments on destruction of different parts of the brain and the retention of learning, memory, and all the rest of it?
DR. KÖHLER: I admire the courage with which Dr. McCulloch tries to relate his neurophysiology to facts in psychology. But, when in a skeptical mood, I sometimes feel like criticizing the results. Take the example of visual shapes which, as we all know, are generally recognized in a peripheral position (or in a larger size), even if, heretofore, they have been seen only in foveal projection (or in a smaller size). Dr. McCulloch's explanation of such achievements introduces more histological assumptions ad hoc than seem compatible with usual standards of plausibility. In fact, he does not seem himself to maintain that a real brain functions in this fashion. Why then the elaborate constructions? Most probably the reason is that the atomistic character of Dr. McCulloch's neurophysiology prevents any direct approach to relationally determined facts such as visual shapes. The difficulty seems to be strongly felt, and special sets of neuron connections are now being constructed which merely serve to remove the difficulties caused by the main atomistic premise. Would it not be simpler never to make this atomistic assumption? If we think of cortical function in terms of continuous field physics rather than of impulses in neurons, the difficulty never arises. The contours of retinal images are projected upon the visual cortex by nerve impulses. Let us assume that here they constitute the boundary conditions of field processes such as electric currents. Under these circumstances, there will be for each set of boundary conditions, that is, for each shape, a particular distribution of a directly interrelated function; in other words, each shape will be cortically represented by a specific process. If the characteristics of such a process remain approximately constant, independently of its location and size, then recognition of a shape in a new place or size offers no problem which is not also present in the recognition of a color in a new place.
Incidentally, it seems to me misleading to assume that the present problem is mainly a problem of recognition, and therefore of memory. When two objects are given simultaneously in different places while the eyes do not move, we can compare these objects, and say whether they have the same shape. Once more the implication is that visual shapes are associated with specific processes.
Occasionally, I am afraid, Dr. McCulloch uses psychological terms in a strangely diluted sense. In fact, sometimes little is left of what they actually mean in psychology. But the change is never mentioned. People will therefore tend to believe that, when such terms are now being related to neurophysiological hypotheses, it is their real psychological contents which are given a physiological interpretation. They will not notice that the essential characteristics of the facts in question are tacitly being ignored. I have an uneasy feeling that this may happen even to the theorist himself. Thus Dr. McCulloch likes to call a nerve impulse a “proposition.” Moreover, he says that the occurrence of a given nerve impulse “implies” the occurrence of preceding impulses (in other neurons), by which the given impulse has been started. But, typically, a proposition is concerned with a relation between certain terms, whatever the relation may be in individual instances. A cortical situation would therefore correspond to a proposition if in this situation the cortical counterparts of two terms were functionally related in one specific fashion or another. A nerve impulse does not in this sense relate two terms to each, other. At least in Dr. McCulloch's neurophysiology, a nerve impulse seems to be a particularly lonely event. How, then, can a nerve impulse represent a proposition? Some discussions of nerve impulses and of their equivalence to facts in psychology make me feel that, inadvertently, an extremely learned histologist and neurophysiologist is tacitly supposed to watch the human brain continually, and that this expert always knows how impulses must be interpreted in psychological terms. He probably tells the owner of the brain what psychological facts he must have when impulses travel in this or that part of the cortical machine. For without this help, what could induce a person to think of a specific proposition, that is, a particular relation between two terms, when an impulse travels in a certain fiber? Since, actually, no such expert is available, the characteristic forms of the various psychological facts must be directly given by the functional characteristics of corresponding cortical processes. But, to repeat, if this is the case, it cannot be nerve impulses which give propositions their relational character. For they have no such character themselves. For the same reason, there can be no connection between the psychological experience that one fact implies another fact and the behavior of a nerve impulse. A present impulse implies, say, preceding impulses in other neurons (McCulloch's example) only in the mind of a neurophysiologist who knows what must have happened a moment ago at a certain synapse. As the present impulse travels along its fiber, it knows nothing of preceding impulses.
For a moment, I must come back to a criticism to which I have referred once before. It must be a hard task to give psychological facts interpretations in terms of nerve impulses. For when this task arises, and is apparently accepted, the theorists soon forget what they must now be expected to do, and turn to other problems which are only indirectly connected with the original problems. Invariably, such substitute problems are more accessible to explanations in terms of nerve impulses. On the other hand, since they are somehow related to the problems which were actually to be solved, psychological concepts which are essential in the latter will naturally also be mentioned when the substitutes are being discussed. Thus, if interpretations in terms of nerve impulses seem to work in the case of the substitutes, both the theorists and others will easily believe that actually the original problems have been solved. For this is what the theorists had promised to achieve.
Take “having a goal” as an example. Before we realize what is happening, the task of explaining this psychological fact in terms of nerve impulses has been replaced by another task: Once a person has a goal, how is the goal actually reached? Naturally, if this is done by overt action, both centrifugal and centripetal nerve impulses will play an important role in the process. It is also a most sensible suggestion that the action is steered in the right direction by negative feedback. But do we learn in this fashion what “having a goal” is in terms of nerve impulses? Plainly, we do not. Nonetheless, we may be so strongly impressed by what seems to have been achieved that we forget what had to be achieved. Of course, it must be difficult to understand “having a goal” as a matter of nerve impulses. “Having a goal” is again a relational situation. When a person has a goal, his self (in a purely empirical sense) is dynamically related to a certain object, and therefore, probably, the neural counterpart of the self to the counterpart of the object. Moreover, the nature of the relation depends entirely upon the perceived characteristics of the object and the state of the self at the given time. The theorists themselves seem to doubt whether interrelations of this kind can be mediated by nerve impulses, which are described as atomic events par excellence. Otherwise, why should the theorists prefer to discuss something else, namely, goal-directed action? And yet, “having a goal” is a problem which must be handled quite apart from overt action in reaching the goal. For people often have goals while they do not yet know how these goals can possibly be reached. It also seems probable that a really adequate interpretation of “reaching a goal” presupposes a correct interpretation of “having a goal.” As a goal is being reached, the dynamic relation between the self and the goal, which seems to represent a store of energy, is gradually being changed—until eventually, when the goal has been reached, this energy is spent. I have a suspicion that the negative feedback involved in the change refers to the store of energy implicit in “having a goal.” But, of course, this again is thinking in terms of field physics.
I will remark only in passing that the substitution of one problem for another occurs also in Dr. McCulloch's treatment of “value.” He does not give us a theory of value in terms of nerve impulses. But values may conflict, just as many other things may conflict, and then the question arises which value will win in a given conflict; that is, which is the stronger value. It is this question with which he prefers to deal. But since the same question may be asked with regard to many facts which are not at all values, we have obviously once more lost our way. We may easily believe that we are actually dealing with the problem of value as such; and this belief will be strengthened by the fact that, in formulating our new problem, we may still mention the concept “value.” But the problem what value means in terms of nerve impulses has in the meantime been forgotten.
Quite probably, Dr. McCulloch will not be impressed by these arguments. He may feel that I am accepting certain premises of which he does not approve. First of all, he is likely to say that the structural characteristics of cortical processes need not agree with the structures of corresponding psychological facts. Actually, he has just told us that the hypothetical cortical counterpart of an idea must fulfill only one condition: It must always occur when the idea occurs. More specifically, he has once said that the cortical counterpart of a square may be a “four-spoked form, not at all like a square.” I cannot agree with this statement for the following reason. It would not be difficult to give subjects a series of tests in which they would have to respond to one structural characteristic of a square after another in overt action. Under these circumstances, the form of their actions would directly follow from the corresponding structural properties of the square. Their actions would prove, for instance, that their square has four straight sides, that pairs of these sides are parallel, that the angles have all the same size, and so forth. From the point of view of natural science, how can this happen if the cortical counterpart of the square has no corresponding characteristics? Does Dr. McCulloch suggest that such characteristics exist only in the square as an “apparition” (his term), that is, as a mental fact, and that, quite apart from the cortical situation, this mental fact as such determines what the subjects are doing? There is an old name for this view. It is called dualism. I find it hard to believe that dualism appeals to Dr. McCulloch. But in this connection he does argue as though he were a dualist.
If we consider how the visual square (the apparition) comes into existence, we meet with the same difficulty. How can a cortical process such as that of a square give rise to an apparition with certain structural characteristics, if these characteristics are not present in the process itself? According to Dr. McCulloch, this is actually the case. But if we follow the example of physics, we shall hesitate to accept his view. In physics, the structural characteristics of a state of affairs are given by the structural properties of the factors which determine that state of affairs. The magnetic field around a long conductor with circular cross section obviously describes circles; the electrostatic field around a charged sphere is symmetrical with regard to the center of the sphere, and so forth. Situations in physics which depend upon the spatial distribution of given conditions never have more, and more specific, structural characteristics than are contained in the conditions. To be sure, this rule holds only so long as the medium in which a physical situation develops is homogeneous, that is, devoid of special conditions of its own. For instance, the field around a charged sphere will no longer be symmetrical about the center if the environment contains various dielectrics in an arbitrary arrangement.
If we apply this lesson to the way in which the cortical counterpart of a square gives rise to this square, we must choose between two possibilities. Either the structural characteristics of the visual square are fully determined by its cortical counterpart. Then this cortical process must have the structural characteristics of the square. Or we assume that the visual square has structural characteristics of its own which are not present in the cortical process. Then the world of apparitions, the psychological world, constitutes a particular medium with special determining conditions, quite apart from cortical conditions; and it is these conditions in the mental world which add the structural characteristics not contained in the cortical process. The second alternative is, of course, again tantamount to dualism. It seems that if we do not want to be dualists we must accept psychophysical isomorphism.
DR. BROSIN: May I again beg your indulgence and give you a full opportunity to continue later? Dr. Lashley.
DB. LASHLEY: I am very much in sympathy with the type of development represented in the last two papers. I think any understanding of the nervous system we may acquire must be developed within the framework of our knowledge of the activities of the individual neuron. There may be additional factors introduced by combinations of which we know little or nothing at present, but the general principles seem to me to be fundamentally correct. At the present time, however, such a formulation involves a very great oversimplification of the problems. The behavior which is explained is behavior which never occurs in the intact organism. It is an hypothetical behavior derived from the assumptions of the system rather than a description of, observed phenomena. A visual object maintains its continuity in spite of constant fluctuations in the position of the eyes and shifts in its position on the retina. By a series of special assumptions concerning neural organization this phenomenon of stimulus equivalence can be accounted for in terms of impulse switching. But in recognition of the visual object it makes little difference whether the image of the whole object falls upon the retina. If only part is seen at any one time, the entire form is rapidly reconstructed from the series of images of parts. The temporal sequence of part figures is combined with the spatial orientation of eye movements to give spatial continuity to the whole. This phenomenon requires a new set of assumptions to make the theory of impulse switching applicable. I somewhat question the utility of a theory which has to be revised to fit each special case.
In its present form the theory of impulse switching involves, I believe, assumptions concerning the accuracy and uniformity of neuronic structure which are not justified by the facts. We have been studying individual variations in the number and arrangement of neurons in the cerebral cortex. We find a wide range of individual differences in cell number and size in corresponding areas of different brains of animals which are grossly indistinguishable in behavior. Two brains may differ by as much as 50 per cent in the number of neurons in the temporal lobe or by 100 per cent in the average size of cells in the superior frontal convolution. A given type of cell may be present or absent from the auditory cortex or operculum. Yet the fundamental behavioral activities of these animals are the same. The anatomic variability is so great as to preclude, I believe, any theory which assumes regularity and precision of anatomic arrangement. Such facts lead me to believe that theories of neuron interaction must be couched, not in terms of the activity of individual cells but in terms of mass relations among the cells. Even the simplest bit of behavior requires the integrated action of millions of neurons; the activity of any single neuron can have little influence on the whole, just as the path of an individual molecule of a gas has little influence on the gas pressure. It is questionable whether specific instances of behavior can ever be dealt with in terms of the activity of individual neurons; the complexity is too great. We shall probably have to use a different kind of model, a model which can be explained in principle by individual neuron action but which involves a somewhat different set of concepts and laws of action. These laws may eventually be derived from study of the individual neuron when those properties are directly observed. At present, however, many of the properties ascribed to the individual neuron are inferred from the activities of neuron masses, and explanations based on such inferred properties are circular and, perhaps, spurious.
Some of the specific hypotheses which have been formulated by Dr. McCulloch seem to me to meet with serious difficulties. He has suggested a reverberatory system between the striate cortex, the suppressor band of area 19, the prestriate region, and the thalamus. I have just removed the prestriate region (including areas 18 and 19) from a series of monkeys and also the frontal eye fields (another suppressor area) singly and in combination. I have been able to detect no visual disturbances whatever following the operations. In no case have we been able to detect significant perceptual disturbances after removal of suppressor areas or of supposed sensory associative areas in monkeys. The specific hypotheses which Dr. McCulloch has suggested for the action of the visual and auditory analyzers imply a definite spatial position of the analyzing mechanisms. Experimentally they are not there.
This leads to the general problem raised by most of the experimental studies of effects of cerebral lesions. Limited lesions or interruptions of transcortical connections produce few or no symptoms. Behavior seems not to depend upon any localized conducting pathways within the cortex. Habits are not stored in any limited area. Such facts point to the conclusion that there is multiple representation of every function. I see no other way of meeting this difficulty except by assuming some sort of reduplicated network of equivalent functional circuits. In other words, we cannot deal with individual conditioned reflex arcs but only with a multiplicity of interacting circuits whose excitatory effects can be transmitted around various types of cortical interruption.
One other point, in relation to the problems of memory raised by Dr. von Neumann: For memory there is the same problem of equivalence as for transneural conduction. I have found, for example, that one sixtieth of the visual cortex of the rat will mediate visual memories and it may be any sixtieth, provided it includes part of the central projection field. Here, again, there must be some sort of multiple representation. The memory is not stored in a single locus.
Now consider the nature of a memory. It is not a single item which can be filed in a single neuron or reverberatory circuit. It is always the capacity to reproduce a series of events, to reproduce a complex sensory pattern or a series of motor activities. Such neural events involve the activity of millions of cells. I have come to believe that almost every nerve cell in the cerebral cortex may be excited in every activity. I shall give some quantitative evidence of this tomorrow. Differential behavior is determined by the combinations of cells acting together rather than by cells which participate only in particular bits of behavior. The same neurons which maintain the memory traces and participate in the revival of a memory are also involved, in different combinations, in thousands of other memories and acts. The memory trace is the capacity of many neurons to work together in certain permutations. In a system of interconnected neurons the number of possible permutations may greatly exceed the number of switching mechanisms. Perhaps this answers Dr. von Neumann's difficulty with regard to number of elements. It is also an argument against the dynamic as opposed to static character of the memory trace.
DR. BROSIN: Dr. Weiss, have you anything to add?
DR. WEISS: Much of my comment had better be left for a later part of the symposium. For the present I want to point out that we are actually dealing here with two different problems. Namely, first a statistical consideration, as it were, of whether or not the number of elements present in the nervous system and their various interrelations is sufficient to account for the number and variety of things it can do. We realize, and this gives us intellectual comfort, that the number of possible constellations is large enough to allow for the observed variety of behavior. This statement, to quote McCulloch, would be merely tautological. As a biologist, I am more interested in the second problem, and that is the precise pathways and chains of processes through which, out of the infinite variety of possibilities, just the appropriate sequence and selection are activated which lead to a given appropriate organized response. And if we deal with these mechanisms not as abstract categories, but in concrete terms, then I see some serious and realistic difficulties arising for any theory of nervous networks that requires the amount of precision postulated in the schemes here presented. The study of the developed nervous system, with which the anatomists, physiologists, and psychologists are usually working, suggests a high degree of precision in the arrangement of the constituent elements, but it must be realized that this impression is illusory. The organizational stability of a performance of the nervous system is much greater than the precision of the underlying structural apparatus.
I have referred above to the persistence of the response after experimental or pathological interference with the anatomical substrata of nervous activity, but want to point now to an even more impressive fact, namely, the great variability in the degree of precision of the anatomical networks in the course of development. The fact is that we frequently suspect a given neuronal precision setup as being relevant for a particular neural function, but often find that in an earlier stage of development this function will be performed in essentially the same way without that particular structural precision scheme having even developed as yet. In general, many a condition which we would think essential from the study of the developed nervous system loses pertinence when studies of earlier stages show that things work very much the same even in its absence. This must be emphasized particularly in connection with the present discussion of the relation between input and output of the nervous system. It is a fact that most of the basic motor patterns of behavior are developed within the nervous system by virtue of the laws of its own embryonic differentiation without the aid of, and prior to the appearance of, a sensory input from the outside world. The basic configuration of the motor patterns, therefore, cannot possibly be a direct product of the patterns of the sensory input. A study of the development of the nervous system and of behavior forces us to consider the output of the nervous system and its patterns as primarily preformed within the nervous system and ready for use, requiring the sensory input for release, facilitation, and modification, but not for its primary shaping.
This brings us to the fundamental alternative, to which I think Dr. Gerard has likewise referred, of whether the central nervous system is merely a clearing house for input-to-output messages, or whether it generates activities of its own and has patterns of activities of its own, the elements of which are not pieced together by, and reflections of, the sensory input. Dr. Köhler has likewise touched on this fundamental difference in the interpretation of the realities of the nervous system. No theory of the nervous system can claim to represent the facts if it ignores the central autonomy of the basic patterns of motor performance. This autonomy impresses us not only in the studies of the development, but also in studies on reconstitution after injury of the nervous system, which touches on a question Dr. von Neumann has raised with regard to learning. This is the question of whether learning implies a complete reorganization of the nervous network with a resetting of relations among individual neurons, or the acquisition of a new performance, which will merely supersede, rather than replace, the older performance. This question can be crucially studied by disarranging the peripheral motor apparatus by crossing tendons or nerves, and thus rendering the original impulse patterns inadequate for the performance of a given act. Experiments by my former student, Sperry, have shown that rats cannot relearn their motor coordination to meet such new situations. Studies we have made on patients with transplanted tendons after partial infantile paralysis show that they can learn to use the transplanted muscle in its new function, but precise electro-myographic records show that the muscle is apt to lapse back into its innate phase of activity, thus proving that the learning act does not dissolve the original patterns of motor organization. Evidence of this kind demonstrates clearly that the act of learning does not consist of merely a recombination of individual neuronal elements. On the basis of all existing evidence, the nervous system must not be conceived of as a network of monotonic elements, but as a hierarchical system in which groups of neuronal complexes of different kinds are acting as units, the properties of which determine the configuration of the output pattern. Some of these higher units are rigidly fixed in their functions, others are modifiable by experience. I fail to see this hierarchical principle duly reflected in the theory of a monotonic network of units such as has been discussed in this session. ...
DR. WEISS:2 Let me start the present discussion by stressing the great change in our thinking which the facts we have been hearing mark over the state of affairs twenty or twenty-five years ago, when the hard, fast, and precise one-, two-, or three-neuron arcs were viewed as the keys to the understanding of nervous activity. In the meantime we have come to recognize the tremendous complication of the system. This development ought to caution us against trying again to draw a final picture with the material available to our present fragmentary state of knowledge. We ought to expect that in another twenty-five years additional essential parameters will have been discovered. We have been impressed here with the tremendous complication of the nervous network as far as the shape, distribution, and ramification of the individual elements is concerned. To this intricacy of spatial patterns, I am sure, will have to be added, as another parameter determining nervous activity, specific constitutional differences of biochemical kind among the elements, as well as specific states of activity of larger collective units based on those chemical differences. If I may briefly elaborate this concept, we have been treating the nervous system as essentially composed of units of a single kind. Dr. Lorente de Nó has emphasized the existence of a great variety of what he calls “types” of which only three or four have thus far been studied, while thousands of others are still to be explored.
But what is really meant by a cell type? Does the term merely signify a characteristic shape of the cell body, distribution of the processes, and geometric configuration of the arborizations? Or does the term mean to imply the existence of different cell species with distinctive chemical properties? Is the protoplasm of all nerve cells the same, merely cast into different forms and arrangements? Or do these different types of nerve cells have distinctive chemical characteristics that are instrumental in the operation of the nervous machine? Evidence is increasing that we shall have to adopt the latter view. The variety discernible under the microscope is merely one of the relevant differentials among nerve cells. If we had only microscopic pictures of different glandular cells, we might consider them to be of one kind, but we know that their products and their production machineries differ fundamentally in chemical regards in spite of their similar appearances. Likewise the microscopic resemblance of all nerve cells is no evidence of their similarity chemically. There are two problems to be faced. First, are there significant chemical differences among neurons? Second, if so, what is their role in the operation of the nervous system? As for the existence of chemical differences, the evidence seems clear. Pharmacology has demonstrated that different drugs have a selective affinity for different types of nerve cells. Such different response to chemicals can only be based on a fundamental difference in the chemical constitution of the responding neurons. Similarly, when we classify neurons as cholinergic or adrenergic, that is, as capable of producing different types of substances, we admit that the fundamental constitution of their protoplasms must be different. There is evidence that different types of nerve fibers, such as sensory and motor fibers, and perhaps even different subclasses of fibers within each category, follow different routes in their primary outgrowth during development. This again expresses constitutional chemical differences between those types, endowing them with selective chemical affinities by which they can follow different tracts. Again in establishing peripheral connections with effector or receptor organs, and similarly perhaps between different intracentral junctions, selective discriminatory faculties come into play which can only be conceived of in terms of specific chemical sensitivities based on the chemical organization of the respective units. The existence of chemical differentials between neurons has thus been established beyond doubt. The question then arises whether they may be dismissed as simply instruments in the structural development of the nervous system, which leave their residues in the adult body but are irrelevant as far as the operation of the nervous system is concerned, or whether such chemical differentials are significant factors in nervous activity. That is, do we have to add chemical parameters as determiners of nervous activity to the known parameters of the conduction process, such as time sequence, frequencies, thresholds, synchronism, and other space-time characteristics based on the electric activity of a characteristically distributed network? I submit that we must answer in the affirmative, that we must recognize qualitative differences among neurons as decisive in the making and breaking of functional connections, and that chemical conformance or nonconformance between elements may decide whether a synaptic junction will be passable or impassable.
This brings into view a whole new field of possibilities, which it seems can derive substantial support from general biological experience. It may be appropriate to point out here this more general biological background. As I have outlined in the past several years, many relations between contiguous cells are explicable in terms of the conformance or non-conformance between the configurations of the molecular surface populations occupying the contact surfaces. That is, the behavior of two contiguous cells will differ fundamentally, depending on whether or not molecular key species on either side of the contact surface interlock by virtue of their steric relations. We may visualize these relations in the old key-lock picture of Ehrlich in the modern version given to it in Professor Pauling's theories or in any similar fashion. Such relations are the basis of enzyme reactions, of serological reactions, presumably of growth and reproduction, and as I have tried to show, of intercellular relations. If specific intermolecular bonds along the surface of cells are the underlying mechanism of cellular affinity in general, then it would be quite plausible to assume that the making and breaking of connections between neurons might likewise depend on the conformance of the molecular patterns along their surfaces. That is, given a stationary microscopic network of connections, the impulse could pass only from one element to another if the molecular populations along the barrier were of matched character. Since, as I have indicated in another place, the molecular border population of the cell will vary, depending on the physiological state of that cell, a cell with a given chemical constitution can assume a considerable variety of surface states and accordingly play different parts in intercellular and supercellular activities. It is possible that the proper setting of these border conditions among contiguous elements forms a process quite distinct from and preparatory to the conduction process, and if this is true, then you realize that we have hitherto omitted a very significant coordinating principle from our nervous theories. To the parameters of space and time may have to be added those of chemical conformance and non-conformance. In this connection I should like to call your attention to the fact that in this morning's discussion Dr. McCulloch has illustrated his theory solely by examples taken from those sensory fields which have spatial organization, that is, the optical, tactile, and acoustic fields. In these cases, it is relatively easy to make a case for the patterning of the response by switch-work systems.
However, when we turn to non-spatial qualities of sensation, such as color, taste, or smell, the resolution of these qualities into spatial distributions certainly presents difficulties that cannot be glossed over. Let us consider for instance the experiments of Curt Richter in which rats proved capable of selecting among a variety of foods the one most appropriate to an artificially produced deficiency state of their bodies. Or take the ability of a parasite to recognize very specifically its prospective host by certain chemical cues of the latter. It seems to me impossible to account for these facts otherwise than by conceding to the chemical specificity of the neurons a decisive role in the operation of the system. If in the future we will focus attention on the chemical differentiations among neurons as the presumable mechanism of establishing and breaking intracentral functional relations, we shall undoubtedly find more supporting evidence for this concept. Of course, the microscope will not help us in this task except in so far as a specific differential stain is frequently a safe indicator of biochemical diversity among cells.
DR. BROSIN: This striking development certainly deserves more discussion. Perhaps we can persuade Professor Pauling to comment on it later in the week.
DR. GERARD: Since I have been insisting for years that there are chemical differences between neurons in the different systems, I am obviously, in general, in complete sympathy with the approach which Dr. Weiss has just urged, and I think it might be stimulating to apply it to the question of memory. If that should depend on the passage of impulses across synapses, thereby leaving them altered, might not the alteration, in fact, be a kind of molecular reorganization of the surfaces at the junction, of the sort suggested? I am not inclined to be too optimistic about this extra freedom helping us greatly at the present time, at the level of analysis which is possible with the present procedures—even the chemical ones. Unless one could progress by logic, there is the problem of getting antigens out; and it would be pretty terrific if evidence were obtained that neurons can interact only if they are of the appropriate chemical specificity to match with each other. I think this can perhaps be dismissed at the present time on three grounds. One, although in normal function many neurons are functionally inaccessible to each other, with appropriate drugs they do become accessible to each other—as in strychnine poisoning. Two, even in the normal reflex chains, cholinergic preganglionic fibers act on adrenergic postganglionic fibers. And, third, even within a single neuron itself, there is unquestionable chemical specification, the metabolic reactions of nerve cells being different from those of the nerve fibers which come from the nerve cells. One prefers glucose as fuel and the other glycogen. Cortex is inhibited differentially more by iodo-acetic acid and medulla by malonate. And even in degenerative diseases, the absence of one of the vitamin B components leads to degeneration in the cell bodies of a sensory neuron group, the fiber degenerating only secondarily; and absence of another of the B components leads to degeneration of the fibers, with the cell bodies only secondarily showing disturbance. Therefore, I doubt if we can make too much of chemical specification as a basis for function at the present time, however valuable it may be in interpreting time and space sequences in the development of brain, nerve, muscle, and the like.
DR. KLÜVER: It is of interest that some of the older neuroanatomists, more than thirty years ago, insisted on the importance of chemical differences in the nervous system. In fact, they provided data in support of a chemical topography by systematically studying the absence or presence of various substances in different regions of the nervous system or in different types of cells and fibers. Although the methods used may seem crude in the days of modern cytochemistry there is no doubt that the importance of chemical differences for the study of normal and pathological functions was recognized long ago. When I once discussed these matters with my friend, Dr. Polyak, I ventured the opinion that each nucleus in the brain stem differs chemically from every other nucleus. He not only agreed, but insisted that each neuron differs chemically from every other neuron. When I asked him for his reasons he replied, “All neurons have different shapes.”
DR. WEISS: My reference to chemical differences among nerve cells has not been altogether understood. The gross detectable differences which I mentioned and to which may be added those of preferences for nutrient substrates and other metabolic differences, I have used only as crude indicators of the existence of differentials; but the relevant differences which I think are instrumental in the operation of the nervous system are of much subtler character. We cannot even be sure whether these differences concern the chemical composition of the cell or merely the relative distribution of the chemical key compounds present in the cells, since what determines the behavior of a cell under given conditions is only the particular segment of the molecular population that happens to occupy the surface and thereby controls what is going into and out from the cell as well as what impulse relations the cell will establish with its neighbors. Such subtle differences of organization will hardly ever be detected by grinding the cell up and making chemical bulk determinations. The main purpose of my remark was to call attention to the presence of such subtle chemical relations as instrumentalities in biological relations in general and to call for more intensified study in the case of the nervous system. I may add another biological comment, returning to Dr. Lorente de Nó's remark that his identification of cell types referred to characters of shape and spatial organization. The study of developmental mechanics has shown that the shape of a cell is only an expression of responses of the cell to its environment in the course of its development. Thus if two cells in an otherwise identical environment assume different shapes, this can generally be considered an expression of underlying differences in the response mechanisms of these cells, or in other words, of differentials in their physiochemical make-up. This is further evidence of the constitutional diversity of the cellular elements in the nervous system.
DR. BROSIN: I have asked Dr. Wiersma of the Biology Department here to tell us of some of his work related to the problem under discussion. Dr. Wiersma.
DR. WIERSMA: I'm glad to have this opportunity to bring before you some work I have done with the synapses of the crayfish central nervous system and at the same time to put some questions to Dr. Lorente de Nó that occur to me concerning the influence of the structure of synapses on transmission. The central nervous system of the crayfish with which I have been working is a rather fortunate contribution of nature, in that it offers, for one thing, a postganglionic fiber which can be stimulated by different preganglionic elements, three of which can be isolated and stimulated as single fibers. These elements are the giant fibers, which run through the whole length of the central nervous system. On stimulation with a single shock, the resulting impulse in any of the giant fibers causes a discharge in a motor root of each of the abdominal ganglia, and the animal flaps its tail. That synaptic transmission is involved can be shown by the fact that stimulation of the motor root does not give rise to activity of the giant fibers. The synapses between the giant fibers and the fiber in the root which discharges have been described by Johnson (1924). If one makes a crosssection of the cord, one will find that the motor root fiber, which itself is of sizable diameter in the root, arises from a ganglion cell on the opposite side. It travels straight up and makes first contact with the heterolateral medial, then crosses over to the homolateral medial, next lays itself against the homolateral lateral giant fiber, and then enters the root. The places of contact are most likely the places where transmission occurs. We find that indeed, by leading off from the root, the root potential follows closest after the impulse in the homolateral lateral fiber, somewhat later after the homolateral medial impulse arrives, and the greatest delay occurs when the heterolateral medial is the activating fiber. Stimulation of the heterolateral lateral fiber always results in excitation of the homolateral lateral, since these two fibers are connected in each ganglion by a synapse, which has some special properties of its own which time does not permit me to describe.
Now, it is possible to bring the preparation into a state in which a single impulse in any of the giant fibers will no longer result in a root potential. Under these circumstances, we will find that if we put two impulses in any one synapse, a root potential will result for time intervals up to about 15 milliseconds. This is, of course, a fairly long time for synaptic summation. However, there is a stranger phenomenon: We can, when the preparation is in this state, combine the stimulation of any two giant fibers and obtain within certain time limits, root potentials. Another remarkable fact here is that simultaneous stimulation of the postganglionic fiber at two synapses does not in general result in a root potential, but that the two preganglionic impulses have to arrive with at least a certain interval. The period during which no root potential is obtained I have called the inert period. If we combine two neighboring synapses, this period has a length of about 2 milliseconds, whereas if another, inactive synapse intervenes, its duration is about twice as long. At the same time the period during which summation can be obtained is shortened with increased distance between the synapses. In other words we may conclude that a process must spread with a certain speed and diminishing intensity from one synapse to the other.
There is pretty good evidence that excitation indeed takes place at the site of the secondly stimulated synapse and not somewhere between the two synapses. In the case where a single impulse causes a root potential, there is, as mentioned, a difference between the delays, when different giant fibers are stimulated. Similar differences are present when summation is necessary, and the delays obtained in this way correspond best with those of the second fiber stimulated. There must thus be a conduction of some process which causes increased excitability. The nature of this process is uncertain: chemical, electrical, or a combination. From the duration of the inert periods and the distance between the synapses, it is possible roughly to calculate the speed of spread, and this comes out as about one fiftieth of the normal conduction speed of the action potential of the root fiber. There is still a fourth synapse, of which less is known, by which the root fiber may be stimulated. This one can be stimulated by exciting the first root on the same side as the third root, from which the lead is taken. Since the whole first root is stimulated, there may be more than one synaptic connection involved in this case. Nevertheless, in combination with giant fiber stimulation, first root stimulation gives results which fit well into the picture. Thus the inert period is longest when the distance is greatest, which is the case when lateral giant fiber and first root stimulation are combined.
The synapses described previously are by no means the only interesting ones which the crayfish has to offer. There are also the synapses in the lateral giant fibers. These are reminiscent of similar structures in the giant fibers of the earthworm, as worked out among others by Dr. Bullock. These macrosynapses consist of a joining of two nerve elements of the same diameter by a membrane. I have tested this membrane in the lateral fibers mechanically. By pinching the fiber below the membrane the end will swell up quite considerably, but no fluid will go into the anterior fiber. Also, on pulling, the two fibers do not come apart. There is, for all practical purposes, a complete anatomical division between the two nervous elements. Nevertheless conduction takes place in both directions with equal facility. My opinion is that this may be due to the wide contact area which this synapse forms. In the above discussed synapses, the postganglionic fiber is much smaller than the preganglionic and hence a large part of the circumference of the postganglionic fiber is involved in the synapse, but only a limited area of the giant fiber. The latter factor would make it impossible for the action potential of the root fiber to stimulate the giant fiber, and hence one way conduction. I would like very much to know from Dr. Lorente de Nó, whether or not he would make such purely anatomical differences responsible for differences between other synapses.
DR. McCULLOCH: May I ask one question? In this buck-jointed axon, did you say there is or is not any synaptic delay?
DR. WIERSMA: I do know that if there is a synaptic delay it is less than one tenth of a millisecond. One cannot say there is none, because the measurements are not more accurate than this.
DR. LORENTE DE NÓ: Is the mechanism by which you change the state of the ganglia cells rapidity of stimulation?
DR. WIERSMA: Yes, but aging will have a similar effect. Once brought into this state they remain in it for a time. The delay is also not constant in these preparations. The longer you stimulate, for instance with an intermediate frequency of 20 per second, the more the synaptic delay will increase, until the root action potential drops out.
DR. BROSIN: Thank you, Dr. Wiersma. Dr. Lashley?
DR. LASHLEY: I should like to point out the wide ramifications of the work of Dr. Lorente de Nó, work which has been revolutionary in many fields of study of the nervous system and of psychology. In the first decades of this century, the view was widely held (at least among those of us who are somewhat naïve in neurophysiology) that the conduction in the nervous system is always downstream. As a result of that view, in large part, came the development of behaviorism; the denial of imagery (memory images) because there seemed to be no mechanism for central reinstatement of activity, and the development of motor theories of thinking by Watson and others. There seemed to be no mechanism for continuity of thought processes except a chain of circular reflexes involving muscular contraction and the resultant stimulation. Many theories of learning at that time were also very strongly influenced by this conception. The dominance of the belief that the conditioned reflex is the primary form or prototype of the mechanisms of learning derives in large measure from this concept. Dr. Lorente de Nó, in his demonstration of reverberatory circuits, provides a mechanism for central maintenance of activity and so throws an entirely new light upon many of these problems. His studies are now leading to a rather widespread revolution in psychological as well as in neurological concepts. ...
DR. BROSIN: With the thought that we would continue the discussion of the previous speakers in as unified a form as possible this morning, we will pay Dr. McCulloch the most sincere of compliments by working hard at re-examining his propositions. Therefore, continuing what we began earlier, I will call on Dr. Liddell to tell us about the difficulties in methodology of animal behavior as it relates to the neurophysiologist.
DR. LIDDELL: Dr. McCulloch's paper, as the discussion of yesterday demonstrated, was admirably provocative and, in my own case, led to disquieting nocturnal ruminations concerning psychology in its present relation to neurophysiology. May I, therefore, give a sheep's- eye view of the matters under discussion; that is to say, from a strictly pastoral point of view.
In the first place, most of the experimentally observed facts of neural function and the structural details upon which Dr. McCulloch's theoretical formulation is based are derived from mammals simpler than man. Why, then, would it not prove more rewarding to attempt to fit facts derived from systematic experimental observations of the behavior of the simpler mammals—sheep, goat, cat, dog—to the theory rather than select instances of human behavior to support the theory? This can be done and should be done.
I propose Pavlov's method of the conditioned reflex as almost ideal for the purpose. Conditioned reflex study has for too long been misunderstood and for a simple reason. Pavlov's method has yielded a mass of verifiable facts in search of a theory. Dr. McCulloch's admirable theory is in search of facts. Is it not possible that, first, Pavlov and, now, McCulloch suffer from a painful phantom limb? In Pavlov's case the phantom limb was an oversimplified imaginary nervous system which could not support the weight of facts. Perhaps McCulloch's painful phantom is the colorless ghost of real behavior. Let us see how we may relieve McCulloch's suffering.
Gasser has said that the electrophysiologist can answer the question “when?” but not “what?” Here is precisely the advantage of Pavlov's method. The conditioned animal can be trained quite exactly to tell when by salivation, leg flexion, cardiac acceleration, or other physiological indicators. Moreover, we can fairly exactly explore the operational limits of its neural computing machine through discovering which forms of trained anticipation lead to breakdown or experimental neurosis. We can find its most vulnerable spots.
The sheep, goat, or dog, trained to self-imposed restraint, through the use of a comfortable restraining harness, learns to expect electric shock or food, or acid in the mouth, as Pavlov found, at some signals but not at others. Some dogs, but not all, when thoroughly conditioned to a series of positive and negative signals, may develop experimental neurosis when the signals are suddenly reversed, where, now, all food signals mean “no food,” and “no food” signals are all followed by food. We have verified this neurosis-producing procedure in the pig. However, a slight change in Pavlov's method of reversing signals will render it innocuous. If a sheep is trained for several months to expect mild shock on the foreleg following a buzzer sounding for 10 seconds, and no shock following the ringing of a bell for 10 seconds, these signals may be reversed with impunity twice within a single two-hour period by proceeding as follows. The buzzer sounds; the sheep alerts; but just before it is able to raise the foot from the platform, the buzzer ceases and no shock follows. The bell sounds for a few seconds, and shock is given. In very few repetitions of this cut-off procedure, the sheep has correctly adjusted to the reversed signals— flexing the leg at the bell and remaining quiet at the buzzer.
Another simple method of testing the sheep's and the goat's neural computing machine is as follows. A telegraph sounder clicks once a second for 10 seconds and a shock is applied to the foreleg. After 2 minutes, the telegraph sounder clicks again for 10 seconds, followed by shock. Each day, twenty signals of 10 seconds’ duration are followed by shocks, with 2-minute intervals between signals. Neurosis supervenes after about one thousand signals at twenty per day, and persists with no further training for as long as three years. It is characterized by muscular rigidity and slow heart. Where the separation of 10-second signals is by constant 5-, 6-, or 7-minute intervals, neurosis again supervenes; but in sheep and goats, it is a strikingly different neurotic pattern, characterized by diffuse motor activity and rapid, irregular heart. In both types of neurosis, the animal's behavior in the pasture and in the barn is radically changed, with a loss of gregariousness, and with restlessness during the night. If the 10-second signals followed by shock are separated by 1-minute intervals, the animal will submit to as many as forty signal shock combinations per day with no signs of perturbation. Furthermore, shocks spaced at regular intervals of 1 to 7 minutes leave the animal quite undisturbed, although it conditions to the true interval as such, and flexes the limb just before the shock is given.
Work along these lines is technically simple but time consuming. It is, nevertheless, worth the investment of time if it integrates, as I think it does, with the technically more formidable tasks undertaken by Dr. McCulloch in furthering understanding of cerebral mechanisms in behavior.
DR. KLÜVER: Dr. McCulloch told us yesterday morning that he wanted to stay away from the totality of behavior. I am very glad indeed that he didn't make good his threat. As far as I can see, he discussed practically every aspect of behavior, since he considered reflexes, audition, pattern vision, learning, memory, thoughts, ideas, goals, motivation, will, and consciousness.
First, I should like to have some additional information on a few points. You mentioned that stimulation of the primary visual cortex in man elicits the impression of a blurred circle of light whereas stimulation of a spot in the secondary visual cortex leads to the impression of a form. There are reports in the literature indicating that the nature of the photopsiae varies with the type of electrical stimulation employed. It has been reported, for instance, that the photopsiae consist mostly of glowing and colored "roundish forms, disks, or rings" when faradic current is employed, and of stars and ragged forms, such as "pointed sparks," when galvanic current is used. I am wondering whether you care to specify the experimental conditions under which the results you mentioned were obtained. In analyzing the mechanisms of hallucinations, I was interested in the question whether unformed hallucinations ever occur, that is, hallucinations of mere luminosity. It seems that reports on hallucinations never refer to the appearance of visual “dust” or to an undifferentiated luminosity. I am wondering whether you know of any reports referring to unformed hallucinations.
DR. JEFFRESS: Dr. McCulloch also mentioned that the form aroused by stimulating area 18 had a location in space. Was the location relative to the head or to the outside world? You said that it was not relative to the eyes.
DR. MCCUULOCH: It is not located in the visual field, because the patient can turn his eyes toward it. It does not move with the eyes, but stays there so that he can direct his eyes toward it. This is quite different from what is seen when area 17 is stimulated or in entoptic phenomena.
DR. KLÜVER: In connection with your analysis of the reflex turning of the eyes, it seems pertinent to recall Graham Brown's observations on the reflex orientation of the optical axes. He observed, for instance, that the optical axes remain fixed in space if the head of a moderately narcotized monkey is passively moved to one side. This reflex disappears upon removing the superior colliculi or upon increasing the depth of narcosis. It is found to be present in monkeys which are decerebrated at the cephalic boundary of the superior colliculi. Graham Brown was chiefly concerned with determining the influence of cortical activity on this reflex orientation of the optical axes. His work culminated in the suggestion that eye movements obtained from the occipital lobe are equivalent to those which, in normal life, center the visual image of an object fixed in space by bringing it from the peripheral retina to the central field of vision, while movements obtained from the superior frontal convolution correspond to those which, in normal life, keep the optical axes fixed on an object that is moving in space. Graham Brown also suggested that movements obtained from the middle frontal convolution are equivalent to those in normal life where the eyes move at command. I am wondering whether you have any electrophysiological data supporting Graham Brown's idea of such entirely different mechanisms in eye movements.
As regards the general problem of relations between cortical mechanisms and behavior, I am glad, as I said before, that Dr. McCulloch has touched upon so many diverse aspects of behavior. He might be accused, and he has been accused, of talking about behaviors that are very tenuous forms of behavior or even forms of behavior that do not actually occur. Nevertheless, I am hopeful that his analysis may lead to a profitable search for “phantom limbs” and ultimately to full-fledged behavior realities. It is possible that certain forms and consistencies of behavior will be discovered that would escape detection without his kind of analysis.
Finally, I should like to return to the subject of “ideas.” It is not clear to me whether you are content with proposing certain cortical mechanisms that are operative while I am having an idea or whether you really want to write a neurophysiology of logic. You recall that Brickner, some years ago, found that electrical stimulation of a cortical area on the mesial surface of the left hemisphere, at about the junction of Brodmann's areas 6 and 32, resulted in perseveration of speech. When his patient was requested to recite the alphabet she repeated again and again the letter she happened to say at the moment this area was stimulated: A, B, C, D, E, E, E, E, E, F, G, H, I, J, J, J, J, K, L, etc. The repetition continued as long as this area was stimulated and ceased immediately on terminating the stimulation. Perhaps we have here a beautiful demonstration of the workings of a reverberating circuit. A psychologist may have no difficulty in relating circuits of such kind to perseverating ideas. The general point I wish to emphasize is that having an idea, or having even an idea that is perseverating, is different from an idea. Ideas, logical structures, and meanings are in and of a world that is different from the world of physiological and psychological events that occur in the process of having ideas or recognizing and enunciating propositions and meanings, even if such physiological events should represent events in reverberating circuits. I am, therefore, still curious to know whether you want to contribute to a neurophysiology of logic or merely to a neurophysiology that is involved while I am talking about logic.
DR. LINDSLEY: I believe I understood Dr. McCulloch to say, at one point, that he interpreted the alpha rhythm as a possible fluctuation in the excitatory level of the cortex, particularly with reference to the speed of transposition, say from one chord to another in an auditory field or from one pattern to another in the visual field.
DR. McCULLOCH: That is correct.
DR. LINDSLEY: It is a little difficult for me to see how this could be the case inasmuch äs visual stimulation will completely block the alpha rhythm for a period, even after the stimulation has stopped.
Another thing: An experiment not yet reported which was done by Dr. J. R. Smith and myself just before the war (it was interrupted and not continued since—I hope to get back to it) had to do with visual stimulation in relation to modifications of the alpha rhythm. Briefly, it was this. A small spot of light which could be controlled in intensity and in duration, was thrown on the periphery of the retina, and we investigated the effects of its intensity and duration on the blocking of the alpha rhythm. When we got down near to threshold with our stimulation and beyond the subjective threshold, we still got effective blocking of the alpha rhythm. We were recording with our electrodes over areas 18 and 19, so probably very little of area 17 was involved in the case of electrodes on the human head. We were having our subject report after each flash came, and they came at irregular intervals for which he was not warned. When we got to threshold the subject's report would cease. Nevertheless, for a certain region below the subjective threshold, the blocking would still occur. Now, what I would like to have Dr. McCulloch explain, in terms of the McCulloch-Pitts model, is this separation, presumably, of the awareness of the stimulus on the part of the subject and the physiological response that we get. Figure. 1 illustrates the kind of blocking that one gets.
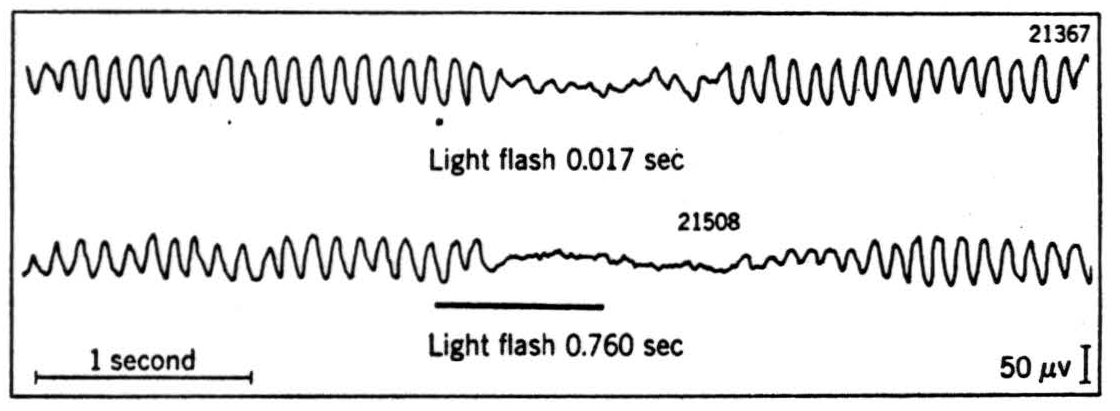
Figure 1. Effect of a light flash upon alpha rhythm.
The illustration does not show what I have just been speaking of —that is, the separation of the awareness or non-awareness of the stimulus on the part of the subject. It simply shows the kind of response we get from a flash of light, in this case, one of which the subject is definitely aware. The duration of the stimulus patch represents the duration of the flash of light. In this illustration, both flashes of light were of the same intensity but of different duration. The point I want to call attention to, however, is that we get this kind of blocking whether the subject is or is not aware of the light, in this region just below the subjective threshold.
DR. McCULLOCH: I wonder if Dr. Halstead would in this connection be good enough to mention his work on the driving of alpha by light. There are probably many people here who don't know the story.
DR. HALSTEAD: If I may postpone it for 30 minutes or so I'll get a slide illustrating the effect.
DR. KLÜVER: Is the duration of the blocking dependent upon the intensity characteristics of the stimulus?
DR. LINDSLEY: It is related to it, but it is related more, perhaps, to the duration of the seen stimulus. In the illustration with the brief flash, after about two-fifths of a second, the blocking begins, and you can see that it persists. Many times you will get really a summation or an after-effect following the cessation of the stimulus. In the second part where the stimulus persists for a longer period of time, you will again see that there is a period following the cessation of the stimulus during which the alpha waves remain blocked and only gradually return, and there you get some of the increased amplitude effect.
DR. KLÜVER: Does the fact that the duration of the blocking is dependent upon the duration or the intensity of the stimulus imply that the awareness or the non-awareness of the stimulus plays no role in influencing the duration of the blocking?
DR. LINDSLEY: I think it is related to both time and intensity.
DR. KLÜVER: Suppose you are able to relate the duration of the blocking to the product of time and intensity. Does this imply that the duration of the blocking is not influenced by awareness or non-awareness on the part of the subject?
DR. LINDSLEY: I don't believe that we have the situation well enough defined to say just what the duration of blocking means as yet. We have in mind further investigation of the problem.
DR. McCULLOCH: Jasper has a good deal of evidence with anxious patients to show that you get disproportionately long disappearances of alpha on very dim illumination even if the patient doesn't see it, and that the duration parallels the severity of the sympathetic overactivity of the patient generally.
DR. WOODBURY: Just to complicate the issue a little further, I would like to mention our experience on the disappearance of the alpha rhythm, depending on whether or not you are getting pattern vision. With electrodes over the occipital region and a towel over the eyes, and the eyes open, we get alpha rhythm with the eyes blank. When we request the subject to focus his eyes on the towel pattern, the alpha rhythm disappears.
DR. BROSIN: Dr. Weiss, do you wish an opportunity to enlarge on this topic?
DR. WEISS: May I mention a few features manifested by nervous activity which any valid theory of the nervous system must duly take into account?
First, the nervous system uses in its operation certain distinct specificities: a specific receptor type and, on the effector side, a specific muscle. Many of you have seen my film on myotypic function in supernumerary limbs. I have demonstrated the same specificity in the proprioceptive system and for the corneal reflex, and Sperry has extended the evidence to the optic field. The existence of such specificities through which discrete peripheral organs and their corresponding central activities are related is an incontrovertible fact. The interpretation, however, is a matter of speculation. Whether we look for spatial order, chronological order, or chemical order as a basis of these specificities, we must always bear in mind that they are distinct, discrete, and show no intergradations. They could be called qualitative differences. Personally I still like to view them as resonance phenomena between complex molecular elements of matching configuration or tuning.
A second condition that any theory of the nervous system must satisfy is that it must be able to explain perception not only in spatially extended fields, such as tactile, acoustic, and optical space— those used by Dr. McCulloch—but must equally well explain recognition of color and chemical character. I submit that this can be done if we concede to the elements of the nervous system the power to recognize chemical constitution by virtue of their own chemical characteristics.
A third condition is that the theory must give equal regard to the role of the molecular processes, the microscopic cellular organizations, and the field properties of the nervous system, all of which seem to be demonstrably involved. How can we reconcile our view that neuronal relations on the molecular level are a coordinating principle with the recognized fact that each act involves vast numbers of neurons whose collective activity behaves in the manner of continuous patterned fields? Perhaps the answer lies in a dualistic concept of nervous activity. We may assume one activity of a conditioning type that sets the stage for a given reaction by determining molecular interneuronal interdigitation, thus linking definite sections of the neuronal network into closed systems, within which, then, a second activity with field character, perhaps merely of electrical order, would operate. Thus by molecular relations, functional subdivisions would be carved out from the total pool of the nervous system, and each such subdivision would then operate field-fashion. Groups of such higher units might then become linked in their activities through correspondences in their respective field activities, and in this fashion we could arrive at a physical counterpart of what we have called the hierarchical principle of central activities. The field character of the activities in neuron pools may force us to extend our current concept of the nervous system in another direction. We may find that the field properties cannot be adequately accounted for by considering only the neuronal elements, that is, by equating the nervous system with the sum total of its neurons. It may become necessary to consider the possible role of the matrix in which the neurons are embedded and learn more of its properties. The environment of the neurons is by no means a physiochemically or anatomically simple system, but is a tissue of complex structural and physiochemical organization. The structural and biological properties of this matrix will of necessity affect the processes in the neuron circuits which are embedded in it and perhaps in counterplay with it. This proposition, perhaps first envisaged by Nissl, certainly deserves further exploration.
A fourth demand on any valid theory of the nervous system is that it must not only apply to the developed operative nervous system, but must explain why the nervous system does not change in its essential functions during growth, although its anatomical substratum is in constant expansion. One must bear in mind that as the minute embryo or fetus or larva grows to adult proportions, the nervous system undergoes enormous alterations in mass, dimensions, interneuronal relations, in the numbers of cells and their sizes. The crucial factors are that the patterns of performance remain relatively stable in spite of these variations of the underlying substrata. New performances may be added, and the precision and speed of older performances may be increased as development goes on, but the basic patterns of coordination remain remarkably constant in spite of the progressive distortion of the underlying anatomical substratum as a result of anatomical growth. This fact brings into sharp relief a question which we have discussed earlier, namely, just how much precision does the nervous system really apply in its performances? If I understand Dr. McCulloch correctly, he tries in his concept to replace mechanical and detailed precision by statistical normalization. I think this is a very fundamental consideration, not only in psychology, but also in biology in general, particularly as regards the processes of development. I wholeheartedly concur in this attitude and hope that it will be more generally adopted. We focus too much attention on precision arrangements among elements, such as a preordained length of a collateral, a rigidly fixed number of collaterals and end feet, a precise diameter of a fiber, and so forth, instead of basing the regularity of a performance on the statistical properties of the collective of units exhibiting it. The regularities observed must be based on the normalization of the statistical distribution of properties in the collective rather than on the minutely precise arrangement of details. This leads us to recognize a certain degree of indeterminacy in biological microevents. This has been true of development in general, and now it seems to be equally true of the operation of the nervous system.
A fifth, and the last condition I want to mention, is one already outlined earlier, namely, that the patterns of motor coordination are not replicas, as it were, of the sensory input patterns. The character and the structure of the motor output is determined by the intrinsic properties of the organized response system and not by the set of stimuli that are fed into it from the environment or the surrounding body. With Dr. Köhler I, too, take exception to calling the light perception, obtained upon pressing on the eyeball, a wrong or false impression in contradistinction to one released by radiant energy. I think it is just as much the real thing as a visual excitation. It is a manifestation of that constitutional ability of the central optical apparatus to generate an activity sensed as light, whether stimulated “adequately” or not. However, whatever the situation in the field of receptors, there is positive evidence that in the motor field, at least, the structural patterns of coordinated response are shaped and differentiated prior to, and without the aid of, a sensory input that could have impressed its structure upon them.
DR. PAULING: This problem of cerebral structure and mechanism may well be related to my field of interest—molecular structure and molecular mechanism—indeed, it must be, as Dr. Weiss has pointed out. I would like to know the order of magnitude of the dimensions of the structures of primary importance in cerebral mechanisms. I suppose (I don't really know anything in this field) that memories—as cerebral processes—involve at first electrical patterns, electrical structure, and that then, after the passage of a suitable period of time, these electrical patterns may be transmuted into material patterns— material structure. And so we ask: How is it possible for this change from electrical structure to material structure to occur, and what is the order of magnitude in size of these material structures? I am not much interested in large structures—structures a hundred angstroms or a thousand angstroms in linear dimensions—but, at least at present, in smaller ones. We can divide things up into different categories of size. First we may consider structures which are, say, 2 to 10 angstroms in dimension—atoms and little groups of atoms, such as the carboxyl group, or an amino-acid residue, that determine mainly the chemical properties of substances. Then we have the next category from about 10 angstroms to, say, 40 angstroms in size. These structures in the main determine, I think, the specific biological and physiological properties of substances. I think that biological specificity is determined almost completely by the size and shape of the outsides of molecules and especially by the phenomenon of complementariness in structure —a sort of lock and key effect. Two large molecules may show a specific relation to one another because of a complementariness in structure. Immuno-chemical experiments have shown that the complementariness must be good to within an angstrom or so, the areas involved being of the order of magnitude of 10 to 20 angstroms square.
Then we have the next category of dimensions, from 40 to 500 or 1000 angstroms, and finally we reach the category of large structures— microscopic structures. This category of structures is extremely important in biology generally; but is it important in a significant way in cerebral processes? I doubt that it is. I think that it is related to cerebral processes, but that it is not important in the sense that cerebral processes involve a change in structure in this size region.
I believe that the specificity of antigens, enzymes, and genes depends on structure in the 10 to 40 angstrom region, that mutation involves a change in structure in this region, and not primarily a change in larger structures. I suggest, then, that the material patterns that are involved in the second stage of learning are the result of changes of this nature, in the 10 to 40 angstrom size category. Now, how could such changes occur as a result of the operation of an electrical pattern that has been induced? Let us assume that there exist molecules, A, that have a definite configuration and that make up part of the nervous system, and also other molecules, A', that are complementary to A, and that are loosely attached to the reticular framework of the brain structure near by. As Tyler has pointed out, there is evidence that mutually complementary molecules exist, perhaps in large numbers, in living organisms. Now, under the influence of an electrical pattern, there may be a change in environment in this region, a change in pH or in ionic concentration, which changes the equilibrium conditions for the molecules A' relative to the reticular portions of matter to which they are attached. This change in environment might permit them to dissociate away, and then to move over and to combine, because of their complementariness, with the molecules A, thus producing a material change which could itself at a later time be reflected in an electrical response.
I suppose all intramolecular forces are electrostatic in origin, inasmuch as they involve electrons and nuclei. Intramolecular forces have been classified as electronic dispersion forces, hydrogen-bond forces, interaction of ionic groups, etc.
DR. KÖHLER: If complementary configurations in your sense occur on the molecular level, and if being complementary or failing to be complementary are facts which may influence function on this level, is it not possible that analogous facts are also important on a macroscopic level?
DR. PAULING: It is very difficult to get a simple chloride ion to attach itself permanently to a structure, because thermal agitation is enough to cause it to dissociate away. You have to have larger aggregates of atoms in order to resist the effect of thermal agitation. Perhaps one of these large aggregates carries a positive charge, and its complementary structure carries a negative charge, and then when combination occurs there is a change in the electrical pattern. In a system where electrolytes are present it is not easy to consider setting up distributions of electrical charge of the order of magnitude of hundreds of angstroms, because neutralization of the charges by the electrolyte occurs, giving a double layer in which the significant dimensions are small. The idea of large-scale electrical distributions hence does not appeal to me very strongly. I prefer to think that some sort of an insulating effect is produced by these molecules, rather than that there is just an electrical pattern.
DR. KÖHLER: I asked my question because, in a discussion of memory, macroscopic field distributions in the cortex are likely to play a major role. I was wondering whether your principle can be applied to such larger configurations.
DR. PAULING: Perhaps it could, since the amount of charge separation required to produce the observed fields is very small. A pattern of loci in which charges are neutralized by complementary structures could be built up.
DR. WEISS: It may be well to point out in this connection that central neuron arrangements, even in the adult nervous system, are much more flexible and variable than we usually admit. Evidence is increasing that the nerve cell bodies react to different functional loads by changes of size. It would seem impossible that a marked increase or decrease in the size of a cell body could occur without the finer terminal connections on the cell body suffering major disruptions. There is, therefore, ample opportunity for rearrangements. We must not think of the microscopic network as something absolutely set and rigid.
DR. PAULING: There is a possibility (though it seems to me to be unlikely) that the whole phenomenon involves changes that are of a still larger dimensional order of magnitude—a thousand, five thousand, ten thousand angstroms. I would be especially interested to know what the characteristic dimensions of the structures involved are.
DR. KÖHLER: I should like to come back to the experiment which Dr. Lindsley has described. The alpha rhythm, he said, can be blocked by a visual stimulus which is too weak to be perceived. This observation seems to be at variance with Professor Adrian's interpretation of the blocking of alpha by visual stimulation. Adrian, it will be remembered, found that presentation of a homogeneous bright field does not disturb the rhythm very much, while even a weak contour may have a strong blocking effect. Since contours attract attention more than homogeneous fields do, he concluded that the rhythm is blocked by attention. But it cannot have been attention which blocked the rhythm in Dr. Lindsley's experiments. A stimulus which is not perceived will be unable to attract attention. I should therefore prefer the assumption that any inhomogeneity in the visual cortex, and in this sense a pattern, is associated with a particular process; that this process is absent, or weaker, when the visual cortex is in a homogeneous state; and that the alpha rhythm is likely to be blocked when such a pattern process is introduced.
DR. GERARD: I am afraid that it isn't that simple, Dr. Köhler. There is another bit of evidence that I am certain most men who have worked with this have experienced themselves. With a little practice, I can look directly at a 100-watt light, which is certainly not a uniform field but is patterned, and, by deliberately paying no attention to it, I can have my alpha waves remain perfectly intact; then with no change except what I can describe in no other way than as directing my attention to the light, have them immediately disappear.
DR. HALSTEAD: We became interested in the relation of vision and alpha rhythm some years ago in a search for neural concomitance of flicker fusion partly as a result of the work on photic driving reported by Adrian and Matthews in 1934. They stimulated man binocularly with intermittent light while recording the electroencephalogram from the occipital areas. The work provides samples of unilateral coupling between the frequency of the intermittent light and the alpha frequency of the EEG. A diffuse (non-patterned) light flash of relatively high intensity was employed to obtain these records—in a conscious individual. About five years ago, my associates and I were able to publish similar records of the driving of brain rhythms, with and without anesthesia, in the Macaque monkey. We obtained a good coincidence over a range from about 18 per second to about 3 1/2 per second. This driving effect can be reproduced with reasonable predictability in several species from rabbit to man. Under the conditions of photic driving the organism becomes literally coupled to the external environment.
We also noted that if trains of flashes, especially in the neighborhood of 6 per second, were suddenly thrown onto the retinae a peculiar hump and spike pattern appeared in some animals at once and lasted from one to two seconds. This pattern is apparently indistinguishable from spontaneous brain rhythms obtained in certain forms of human epilepsy. In recent work by Walters in England, Gastaut in France, and others, petit mal seizures have been induced in man under controlled conditions in convulsive-prone individuals by this technique. The alpha rhythm is abolished for the duration of the seizure and hence may be thought of as being “blocked.” However, our observations along with those of Wang, Bishop, Bartley, Toman, and others indicate clearly that the electrical responses of the normal mammalian brain from rabbit to man tend to follow or to couple with a train of light flashes through the range from about 2 flashes per second to 20 or more flashes per second. Rather than being “blocked,” the most prominent brain rhythm is, under these circumstances, “paced” or “driven” by the photic stimuli. There is some evidence to be found in the literature that suggests that introduction of a contour at the periphery of a flickering field lateralizes the photic driving to the ipsilateral hemisphere. The electrical activity of the contralateral hemisphere is not “blocked,” however; rather it seems to be decoupled from the flickering field.
DR. KOHLER: Can we be quite sure that what is commonly called driving of the alpha rhythm by varying flicker frequencies actually deserves this name? We know that upon sudden illumination of the eye an on-effect can be registered from the visual cortex. Now, when the sudden illumination is repeated, on-effects may be repeated at the same rate. Have we any way of proving that in such experiments the frequency of the alpha rhythm is actually changed by rhythmic stimulation? How does one exclude the possibility that the registered waves are simply on-effects which have the frequency of the flicker?
DR. HALSTEAD: For the conditions of stimulation we employed, it is rather generally agreed that the total on-effect lasts somewhere in the neighborhood of 200 milliseconds. It is conceivable that frequencies up to about 5 per second might be reflected as trains of on-effects. As Bartley and others have shown, however, components of the on-effects are increasingly inhibited as flash frequency increases above this point.
In a favorable preparation and using a glissando technique, one may drive electrical activity continuously and without essential change in wave form from 6 or 7 per second to 20 or more per second.
Another line of evidence is afforded by the enhancement of amplitude of electrical response from the brain during photic driving. This enhancement reaches a maximum around 10 per second in monkey and man. This is generally regarded as the alpha frequency. Toman, studying these relations in man, found it possible to omit a single flash without disrupting the response. We have made similar observations. This would suggest that the centrally induced rhythm may sustain itself briefly without reinforcement.
DR. GERARD: Doesn't the rhythm continue a little while after you stop the flicker?
DR. HALSTEAD: Yes, it does; it may last from 2 to 6 seconds in some preparations.
DR. VON NEUMANN: I don't understand why sometimes even a subliminal signal of light may stop the alpha rhythm, while here it continues in spite of light signals. How are these things compatible?
DR. McCULLOCH: Didn't you get part of an answer to your question in Dr. Gerard's statement concerning the light bulb? Whatever this mechanism is, it is important.
DR. HALSTEAD: I think that there are special circumstances under which the alpha does not block in the presence of light.
DR. KLÜVER: In this connection I should like to ask Dr. Lashley, who has described his scintillating scotomata, whether there are disturbances of the alpha rhythm during the observation of such scotomata. I am also wondering whether such disturbances, if they exist, are related to the frequency of the scintillations.
DR. McCULLOCH: May I answer his question? I have very brilliant scintillating scotomata with hemicranial headache, or migraine. The scotomata have, inside them, an area in which I can see nothing. They begin always right next to the point of fixation, just a whisker to one side of it, and they move out until they have occupied the whole of that field. I already have several times had electrodes well placed in and about the inion and all over the back of my head and there is no change that we can detect. I don't have a good alpha, but there is no change in what alpha I have, and there are no other waves that are not normally there with my eyes open or my eyes closed.
DR. KLÜVER: What is the frequency of the scintillations per second?
DR. McCULLOCH: The thing which I see is composed of small bright dashes and these things don't all come or go in a pulsing fashion, but they twinkle.
DR. KLÜVER: You are apparently not a good case for getting data on the frequency of oscillations. Fröhlich, by the use of a rather ingenious method, found that the frequency of oscillations in his scintillating scotomata was 20 per second.
DR. LINDSLEY: To confuse the issue further, I would like to add a comment. When I was working with infants and young children, where the alpha rhythm is of a different order of frequency, namely, around 3 or 4 per second, I found that the time for delay of the alpha rhythm in blocking is proportionately longer than it is in the adult where the frequency is around 10 per second.
DR. McCULLOCH: There is one more piece of evidence. I don't know whether Mr. Stroud is in the audience, but if he is, I wonder if he would be willing to say something about the rate at which forms can be perceived?
MR. JOHN M. STROUD: Stanford was kind enough to let me come, as a kind of renegade physicist, into the Psychology Department and do a little work on the problem of the frequencies of scansion functions, such as the alpha rhythms (which I considered as the no-load symptom of a scanning function). The hypothesis that I wanted to test led to two experiments that are relevant here.
One was very simple. I had an ordinary cathode-ray oscilloscope and set the observer the task of finding out how fast the sweep was going, by tapping a key which put a little pip on the trace. His task was to learn to tap the key fast enough to make the pip stand still; that was his only task. It showed, incidentally, that human reactions are by no means as unprecise as one might infer from the classical reaction time experiments. Anybody could learn to tap at the rate of 10 cycles per second for one hundred consecutive taps. He could always get his reactions to fall within a band of 20 milliseconds for a prescribed frequency, with errors from one tap to the next of the order of only 2 1/2 milliseconds.
In the second experiment the observer was set the task of matching the brightness of a flickering light by adjusting the brightness of a steady light. The field was a small circle split vertically, with the flickering light on the left and the steady, on the right. The level of illumination was low, and a very dark red light was used. The flicker was rectangular in wave form, and the proportion of the cycle during which the light was on could be controlled by the experimenter. In different runs it was varied from zero to 100 per cent. The experimenter could also vary the repetition rate from about one-half to about 20 per second.
The results varied from subject to subject, but in general showed that for frequencies above about 8 the relation between the proportion of time the flickering light was on, and brightness was linear, with a slope of 1.0. That is when the light is on 100 per cent it is matched by an equal intensity. Call this 100 per cent. Now if the flickering light is on 50 per cent of the cycle, it is matched by a steady light one-half as bright, etc.
If, however, we reduce the repetition rate below 8 per second another phenomenon appears. At 6.4 cycles for some subjects (and at lower frequencies for others) the brightness follows the previous curve for durations down to about 60 per cent. From 60 per cent to 30 per cent it remained constant, and then fell off in a linear fashion with duration.
At a still slower rate (4 per second for many subjects) the brightness dropped very slowly as the duration changed from 100 per cent to about 45 per cent. (The brightness dropped only about 10 per cent in this range.) Then it dropped rapidly as the duration was further reduced. In all cases the data seem to be best fitted by straight lines making sharp corners with one another.
The subjects often reported, where the frequencies were from 4 to 8 and the durations short, that there was a definite light and dark phase and that this was the basis for a phi movement. Most people saw the center line bulge and collapse alternately. I saw a guillotine moving up and down, chopping off the flickering half field. I found that the subject's movement hypotheses were, to a considerable extent, under their control, and that we could have all sorts of hypotheses to cover what occurred at these and at lower frequencies.
DR. McCULLOCH: I would just like to point out that the relations here are just what one would expect from Dr. Halstead's work on the extent to which the alpha can be driven.
MR. STROUD: I know that, but I hesitated, in writing it out formally,3 to call attention to its relationship to the known behavior of alpha although I was familiar with Dr. Halstead's work.
DR. BROSIN: If there are no further questions I will ask Dr. McCulloch to take care of the many that have already been posed him.
DR. McCULLOCH: I would like, if I may, to begin at the other end, so to speak, and thank Dr. Paul Weiss for liberating me. He holds the theory, you see, that the output is in no sense to be determined by the input—it is independent. This suggestion of his makes it very easy to handle the host of questions. I cannot, for the life of me, attempt to answer them question by question. I would much rather try to give what I would call an organized statement about half a dozen major items that are under discussion.
If you will remember, I suggested that we ask the theoretical physicist to account for himself; that is, for organisms like himself, of such a nature that they could produce theoretical physics. Now, in asking that question, I did not ask that he be a psychologist, and I did not ask that he be a philosopher, and, above all, I did not ask that he be a theologian. The entire problem, I am quite sure, will never be manageable if we begin at the most difficult end. I would much rather ask, merely, that it be a theory of the physicist, and my object in doing this is to put the question concerning whether a machine of this order of complexity can state how it works. If it can (and I think it can), then we can build others that can do the same thing. My object in doing this is to keep the issues in a form in which they are manageable from the standpoint of physics and mathematics. I grew up in a medical school where my professor of neuropathology defined the subconscious as a place which is not a place where an idea is when it is not an idea, and my professor of neurology said that he knew that time was in the temporal lobe and space was in the postcentral convolution, but he was not sure where consciousness was.
Now, it is just these kinds of phrases that I would avoid. I don't believe that I brought up the question of consciousness. If I had to, in a medical sense, I would use the word only to say that this patient was or was not conscious, according to whether he could or could not bear witness to what I could also bear witness to. I would, here and now, systematically exclude any such problem. I am, however, full) aware that most of what goes on in me is something of which I am not aware; I only see the end products. I see those only rarely and in funny flashes. Very often I know that I have decided something without even knowing what I have decided. I do not believe that we have any right to ask of whatever it is that is consciousness that it do the chore of alpha waves or something else of that sort. Alpha waves as I see them (and I am coming back to them first and foremost) are a sweep of scansion. That sweep can, to some extent, be stimulus- triggered. It can be pulled down to about half; it can be pushed up to nearly double its idling rate. When we try to push it further, it snaps and gives us double frames or something else of that sort. I think that much is fairly clear. It has this peculiarity regardless of the sensory modality in which one works. One always comes to an ability to follow in the range, from, let's say, five to fifteen—the maximum number of frames that one can take in any sensory modality. The ability to pull out universal seems to be something of the order of ten per second. You can push it up a little or down a little. Therefore, whatever be the method whereby we do pull out universal, it is timed by something that runs around ten per second, and which may be pushed a little one way or the other. This means that all our ideas, insofar as they are timeless, are going to have this peculiar quality of being moments without internal temporal structure, and that our timed affairs, between our ideas, will be between them, and hence of larger temporal units. Now, that is the first point.
Next—as to why alpha rhythm should disappear when the instrument is in use—this has two possible answers. One is that when we go after forms—when we begin to use a device to pull out shapes—we are keying in this mechanism with incoming impulses, and in so doing we upset a sweep in that sense. But it is far more likely that what happens is that when we do use this sweep, the detail (since its voltages are in cells oriented in this same direction) will have its throw in the same direction as the apparent sweep; hence all you will get is that your sweep is passing up and down as a series of voltage fronts and your cells have their major axis in this direction, so they create voltages up and down. Therefore, you would expect that in using this structure you wouldn't see your sweep. In the present discussion I am not particularly interested in which happens.
I should add one more thing before I leave the problem of the alpha, and that is George Bishop's work on the relation of the alpha to an impulse thrown in at various times in the alpha cycle, reported in one of the Cold Spring Harbor Symposia. He showed very clearly that the response that you get in the cortex to an incoming volley is determined in large measure by the positivity or negativity of that cortex—which way up the voltage in it is at the time that the impulses arrive. There is no question but that alpha does affect the appearance of “on” and “off” effects in the superficial layers of the cortex itself, but I am not interested in any one particular mechanism at the moment. I don't care what the mechanism is. What is important to me is that we have some idea, even if it be one very hard to run down, of a process whereby we can get our “stimulus equivalence.” That is the crucial item as far as I am concerned, and I am well aware that in one sense, or modality, it may go one way, and in another in another way, or, in a single modality it may pull out one thing by one method and another thing by another method. This would seem to postulate a theory of a nervous net which has delays and connections in time, and connections in space which are of such a kind that you cannot know beforehand what sort of a thing your invariant will be. It may be a figure of impulses over a single cell, or a series of cells, in time, and it may be over a flock of cells at one time. I have not tried to give a spatial picture of the nervous system or a temporal picture, but a fourfold picture—three dimensions of space and one of time in which that invariance is to be sought. That is the first thing that I want to make clear.
The second thing I want to make clear is much harder to say. I spoke of reverberating memory, and I spoke of reverberating memory to the exclusion of all other memories, not because I believe that all that we remember is so carried, or because I believe that we carry more in reverberation than in any other way, but because anything which can be done by any kind of memory can be done by a reverberating memory, except to go through periods of complete inactivity or complete explosion. As I see it, there are three problems in memory, not one, or three kinds of memory that we are going to have to distinguish always.
First, there is reverberation, and it does work. Second, there is a kind of alteration of the nervous net with use, which I believe is probably crucial in many of the experiments on conditioned behavior. It is obviously so in certain kinds of pathological situations. The third kind of memory, I don't believe could be accounted for by either of these two ways, and it is one with a bottleneck both in putting information in and in taking it out. I am utterly unhappy about where it is. I don't think that we have any better guess today than that it is somewhere in our heads. I see no particular reason for assigning it to neurons instead of to their matrix—the glia. I don't see how we can tell where we have to look as yet, because in many of the experiments in which there are lesions made in brains, we have had large amounts of territory removed. However, usually we fail to destroy most fixed memories; therefore, we cannot today locate the filing cabinets. I think that sooner or later answers to the question of those filing cabinets, or whatever it is on which is printed “photographic records” and what not, will have to be found. The theory that I set up was intended to handle only this question—it was set up to show that organisms or computing machines which digitalized, or quantized, at the level of neurons—at the level of relays—were systems capable of doing certain kinds of things, and rather a large number of things like remembering and computing any computable number.
The question as to whether I am trying to envisage the human nervous system, or that of a particular animal, or whether I'm interested only in computing machines in general is, I think, most fairly answered in this manner. Logical machines, machines for handling logical problems, can be constructed out of almost any material; however, some materials work better than other materials. I do not know out of what materials the human nervous system is constructed, but the principles of design that have to be followed, insofar as they are not matters of the material but matters of the circuit action, will be identical to the extent that the tasks they are competent to perform prescribe their structure. They need be no more, and the theory that I gave (as opposed to the particular hypotheses concerning, let's say, alpha rhythm or visual cortex) is a perfectly general theory. The hypotheses that I made relating to the nervous system are of the kind that will help me to put electrodes in one place to see what is going on in that place and to know what I am looking for, and I expect to hit a snag there very soon. I always have hit snags so far. Now, why have I chosen to quantize in nervous impulses? Well, let's say the human brain is of a general order of complexity of something like 1030 if we think of it in terms of its ultimate particles. One might split this at the level of the atoms, or one might split this at the level of the molecules, or one might split this at the level of the neurons, and so on. The question is: At what level can one split the behavior so as to define a set of units in terms of which to work? And, obviously, the nervous impulse at the level of the neurons is a fairly nice unit for working. It divides the complexity nearly in half. That is why I like to quantize my affairs in the behavior of the neuron and in its properties.
I look on the question of a field somewhat in this manner. In any device (whether we make it a digital device or an analogy device), the ultimate units of our universe are quantized affairs. We use it as an analogy device when we disregard the unitary composition, or definite structure of our elements, and treat them as wholes, which means that we are dealing with their parts, if you will, at best, statistically. We have, with respect to them, thrown away the information that goes into their construction, and have retained only the overall picture of their behavior. Now, I do not like to treat something like the cerebral cortex in this “field” manner. First and foremost, I will admit unquestionably that it has some statistical properties. But what I am looking for is something that will perform a logical task. I like to look for it in a thing that has a grain, and I like to take that grain as my unit, because then I can see what degrees of freedom the system has and to what extent they are bound in any given job of handling information. I believe that there are many things in the nervous system that smear the results. I worked for years on facilitation and extinction. I know that if I stimulate a small area, that there does arise between that active area and any other area, a difference in voltage which persists for a matter of minutes, and so on, with the busy area electrically negative to the depths and to remote regions. I know that if I stimulate this area and it becomes active, there is an increased blood supply; there is a shift to the acid side, and so on; and I know that these will affect, not merely the cells that are active, but (like the direct current that then or there appears) cells that lie in the region, whether or not they have themselves participated, and that this will alter the threshold of those cells.
These happen to be problems that I have worked with and I am working with now, along with the noble metal potentials or oxidation reduction potentials. I am also working on the thermal changes. All these things will smear results. They rob the system of. some of its degrees of freedom without conveying information. They are the kinds of things that would arise if we had to build our electronic devices and keep them in tubs of salt water. I regard them, by and large, for my purposes, as secondary affairs to be worried about, as I worry about the isolation of the paths to ground, so that all of the grounded side of an instrument is free of troublesome voltages or currents. Now, the reason for making this distinction as sharp as possible in the face of plenty of evidence that the smearing occurs, is that we are dealing with something that has a field-like property. My reason for treating it in the manner in which I have treated it is this. It compels one so to state his theory that he can account for these changes by the digital mechanisms; that is, so that he can contrive a digital device to account for each of these. This at once increases his ability to handle his digital machine. It was clearly shown in our first paper, I believe, that these field-like processes—facilitation and extinction—could be handled by sticking in hypothetical nets, because once a system is, in this sense, quantized anywhere, it might as well be quantized everywhere. This keeps the theory clean.
The next question that I want to try to meet is one which is also very tough to state. The cortex, to my mind, is a vast computing machine. I see no reason why a deeper structure may not assign to any particular part of it a particular task today, and tomorrow assign that particular task to some other part of it—except insofar as particular portions of the cortex are stimulus-bound by a path right to them from the periphery. Those paths to it are well known now, and they include a large fraction of the total surface of the total switch board of the cortex. To the extent to which the afferent connections define a portion of the cortex as visual, it must handle something to do with vision. It must handle something to do with touch if it is the part to which touch reports. So, to the extent to which a cortex is so bound back to our sense organs, it cannot be swapped part for part. No lower mechanism can assign to the visual cortex the job of receiving impulses directly from the ears; they only go to the auditory mechanism. But, apart from that, I see no reason why the lower mechanisms may not perfectly well use one part of the cortex today and another part of the cortex tomorrow for the same function. I do doubt that it does this under ordinary circumstances, but I see no reason why, with damage to one part of the cortex, you wouldn't expect it to do just that. As a matter of fact, I think that you would expect it to do that, just as you would expect that when a man's leg is chopped off, or both legs are chopped off, the man learns to get along on his arms. It is a computing machine—it is sitting there—I see no reason why it can't use this part now and that part then.
I would like to come back now, for a moment, to the question of alpha rhythm, this time in cases of hysterical blindness. A hysterically blind man may have an alpha rhythm, and he may have the alpha rhythm with his eyes wide open, and he may have it when you present him with this, that, or the other thing to look at. He doesn't see, of course, but he reacts to it correctly in other ways; however, as far as he is concerned, he doesn't see it, and he may have a very nice alpha all of this time. Yet in the case of the next hysterically blind individual you check, it may be that the minute he opens his eyes, his alpha is gone. Now, I don't think that this hysterical blindness is something that you can pin on any one part of the system. I don't think that when you throw out the works and say, “I shall not see,” that you have to do it in the visual cortex. You may, or you may do it somewhere else, so you may block out alpha or you may fail to block out alpha. And I don't think that when we deal with functions of any great order of complexity, we stand too much of a chance of finding the particular part of the cortex where that is being done, for it can be done now here, now there, except for the sensory projections upon it.
Now, let me contrast this with the answer to the second question that was asked of me, as to why was I so very much worried about the descending systems and so little concerned with the ascending systems. I am worried about the descending systems for this reason. There is no direct path from the cerebral cortex to a motor horn cell known to me. The motor horn cell whose process goes out to the muscle, if it signals to the muscle, has one and only one consequence in the muscle, but, sitting around the motor horn cells themselves, are sacs of internuncials playing around and onto the motor horn cells. To the sacs of the internuncials come stuff, information or impulses, over the dorsal roots, stuff from other parts of the spinal cord, stuff from the brain stem, and so on. The cortex, to a certain extent, tries to take over these other structures by sending signals to them along the way, but its final chore of doing anything in the periphery is achieved only by playing upon the cells which are already going according to ways of their own which are adjusted to the world over hosts of channels. Therefore, what comes out when the cortex starts doing something is by no means simply determined by what goes down from the cortex. That turns out to be actually a small contributory factor to the totality of what is playing upon the motor horn cell. Now, these lower affairs have often very persistent behavior which lasts for a matter of minutes, so that what the cortex does when it starts off something at one time, and then goes to do something else afterwards, is to have to play on systems with enormous (I want to call it inertia) tendencies to overshoot—tendencies to persist in activity. That is why I am worried about the descending system. The ascending system, for the most part, does not have this type of complexity. It has a fairly simple relay system on the way up.
Now, again, I want to make this point very clear. I'm greatly worried about the fact that at the present time we keep trying to assign things to the cerebral cortex that are being done not in it but elsewhere. The reason that we want to assign it to cortex is that if we smash cortex, we don't get it. I feel as though we tried to put the whole organism into the cortex.
The reason I want a clear theory is this. I see people who are insane or who are neurotic. I do not believe that even if you took the best psychoanalysts in the world, and they were a thousand times as numerous as they are, or are likely to become, that there would be enough of them to go around to readjust the neurotics alone, using only auditory afferents to make the adjustments. As a matter of fact, I believe that more and more, we will come to the use of interruptions of the circuit action of the nervous system by drugs, or by electrical stimulation, or by surgery; and the question is, if we want to interrupt something, where is that something happening? How can we interrupt it, and if we do interrupt it, how can we reinstate circuit action that is adaptive? That is why I want this theory, because I hope that it will guide me in placing electrodes so as to find out what is going on where.
Have I left out any crucial questions?
DR. BROSIN: I think that Dr. Gerard is in a position to see, in a judicious way, the trend of the discussion to date, and since the last three speakers have moved the locus of interest a good deal, I wonder if Dr. Gerard would give us a brief summary of what he thinks the vectors have been.
DR. GERARD: I would like to say, first, that I cannot recall attending a meeting of this sort which has gone so happily and so constructively and with such concentrated brilliance. It has been a privilege to be permitted to participate.
The main problem we are attacking is that of correlating behavior with the mechanisms for that behavior. There have been three major approaches by the speakers. There has been an examination of machines, of their basic properties and the logic concerning them, and an heroic attempt made by Dr. von Neumann to see to what extent their general properties apply to those of the nervous system, and to what extent, therefore, they might effectively guide our thinking about the mechanics of the behavior of this most complicated machine. There has been discussion of the nervous system, of the properties of neurons, of their spatial relations, their electrical attributes, and their interactions in groups. And there has been a rational approach, relating various logics to the behavior of the nervous system, which attempted, in a way, to synthesize both of the others. I couldn't help thinking, as Warren McCulloch was making his further statement just now, of how history repeats itself. Some century ago, George Boole became interested in his algebra of logic with the hope that, since the human mind made logic, a study of the laws of logic would necessarily reveal the functions of the human mind and ultimately of the brain. He was perhaps a little more naïve in his time than we are today. He was certainly not aware of, or at least did not give adequate attention to, the non-rational elements of behavior.
Dr. McCulloch just stated that he is interested in devising a theory that would account for the brain as a calculating machine. Several of the questions raised in the discussion have asked about these other aspects. What about the non-rational side? What about the urge for the machine to calculate? What about the drive, so striking in some of the men in this symposium, which, perhaps even more than the elegance of the calculating machine, determines whether or not something worth while comes out of the system in the course of its lifetime? If one wishes to allocate those other aspects to a different box than the cortex, that is perhaps permissible, but it does not resolve the problems that seem to remain.
With calculating, there come inevitably the problems of learning and memory. There has been much discussion of them, and I hesitate to introduce, at this time, any new factual material; but it might be interesting again to make a glissando over the various types of memory that have been suggested. I am always suspicious when I find myself putting things into sharp compartments and saying, “This is this, and that is that.” I don't think nature loves a wall. Some experiments have been reported, by Reynolds particularly, on a type of memory in the nervous system which seems intermediate between the reverberating type and the permanent type. At least, if it be reverberating, it is certainly over far longer times that we ordinarily associate with reverberation. He came to it from problems of pain and relates it to causalgia.
Symmetrical teeth with symmetrical defects in a person's jaw were given dental treatment. On one side the filling was done with great care against trauma and pain, on the other side, with a minimum of this desirable care. In nearly all those patients, severe referred pain developed on the traumatized side. This could be elicited routinely by pricking the opening to the antrum of the maxilla. The sensitivity and referred pain lasted for months, but was immediately and irreversibly abolished by a brief period of conduction block of the nerve from that tooth. Some kind of central overactivity, of inflammation if you will, was able to persist for long periods of time apparently under minimal peripheral reinforcement.
As to the mechanisms for establishing these important behavior attributes, it seems to me that we have wandered in our discussion over a number of hierarchies—the molecular, the neuronal, the cerebral—and the question has arisen whether we need something beyond—the man or the observer—to know what is happening in all of those. This last question can safely be left to the remaining periods of the symposium.
We all agree, I believe, that molecular events are critical in determining neuronal behavior. We all agree that neuronal events are critical in determining the total behavior of the nervous system itself. The discussion has focused mainly on this step. Dr. McCulloch has explored it from the particular viewpoint of the nervous system as a digital machine, functioning exclusively in terms of quanta, of the nerve impulses which are or are not discharged by particular neurons under particular conditions. Others have emphasized other kinds of relationships, of the analogical type. The real question being asked, I believe, is whether, in addition to a rigid hierarchy of interaction from molecular to neuronal to total performance, there can also be a direct influence from the molecular level to the molar level.
This question is also involved in the one of whether neurons can interact with each other to form some sort of total functioning unit by mechanisms in addition to the simple quantal discharge of elements.
At the one extreme, the position could be taken that all that happens in the nervous system depends on digital machines and unitary discharges. At the other extreme, the position could be taken that these are rather trivial in the functioning of the nervous system, having to do with the more routine activities controlling movement and the like; but that all the more complicated functions depend on analogical behavior, involving chemicals, electrical fields, spontaneous changes, which might act in many other ways than as a scansion mechanism. Some might say that these are the power determinants of the more complicated types of behavior. I sincerely doubt, however, that anyone in the room would take either of those extreme positions. Rather, we are all prepared to recognize that both elements enter into the picture. The point of disagreement would certainly be about the relative importance of the various mechanisms and their relative significance in particular aspects of total behavior. That is as far as I should go at this time.
I was a bit shocked by Dr. McCulloch's comment, a moment ago, that all the other phenomena are merely degradations in the functioning of a beautiful digital mechanism. I really don't think that he believes that, and perhaps I misunderstood; for the very alpha rhythm that he uses as his scanning device is non-digital. In any event, the speakers who are to follow will surely emphasize these other aspects, the mass action effects and the potential fields, and we have already had a considerable attention to the chemical aspects, which likewise transcend the simple matter of discharge of impulses along neurons. We can confidently look forward to equally stimulating and provocative periods in the remainder of the symposium.
DR. VON NEUMANN: I would like to bring up something that I consider remarkable and something which I think did not come out very explicitly except in one remark of Dr. McCulloch's, and I would like to emphasize it. I think that in regard to the great flexibility, there are few symptoms—if one thinks of the functioning of the nervous system and compares it with the functioning of very complicated automata—which are very conspicuous in the case of the nervous system and very conspicuously absent in any automaton that we know of, which may be easily related with each other and related with the circumstances which one ought to expect to emerge at this level of complication. It is very obvious that the brain differs from all artificial automata that we know; for instance, in the ability to reconstruct itself (as in the case of mechanical damage). It is always characterized by a very great flexibility in the sense that animals which look reasonably alike and do the same thing, may do it by rather different cerebral mechanisms. Furthermore, though all humans belong to the same category and do the same things, outwardly, in some cases they are using different cerebral mechanisms for the same things, so there seems to be a flexibility of pathways.
There are some indications (I do not wish to go into them in detail now) that some parts of the organism can act antagonistically to each other, and in evolution it sometimes has more the character of a hostile invasion of one region by another than of evolution proper. I believe that these things have something to do with each other and also with the following. The largest automaton which we know how to plan could consist of something of the order of 104 units, and this is already very complicated. By this, I mean the fact that they are still nominally made from blueprints, and, in all cases where an automaton is built, there exists a blueprint for it, and there is somebody around to guarantee that it was built from the blueprint. These things are, however, very marginal. It is probably not true for most automata of this degree of complexity that they are really identical copies of what is on the blueprints. And it is probably no longer even true that there is anybody around who had anything except a somewhat intuitive relation to it. This is increasingly true for complicated machines. No amount of time and skill and reliability in components will change this in an automaton more complicated than this, and they will probably become more and more complicated. There are a hundred thousand to a million parts involved in some of them, and this is still low, compared to the brain. Building will probably be a completely nominal operation, not completely controlled by what one finds in blueprints. It will have a different relation to the object, and by that relation, I mean this. It has already happened (and it is, of course, just by the introduction of automata into mathematics that it begins to happen) that you are no longer thinking about the subject, but thinking about an automaton which would handle the subject. It has already happened in the introduction of mass production into industry that you are no longer producing the product, but you are producing something which will produce the product. The cut is, at present, never quite sharp, and we still maintain some kind of relation with the ultimate thing that we want. Probably the relationship is getting looser. It is not unlikely that if you had to build an automaton now you would plan the automaton, not directly, but on some general principles which concern it, plus a machine which can put these into effect, and will construct the ultimate automaton and do it in such a way that you yourself don't know any more what the automaton will be. You will do it in such a way that all you will know is what you want to do by it, and by what qualitative methods it can be done, and how to make an automaton which can put this into effect. But as to how the primary thing does it, you need not know. You don't need to know what the result of every particular multiplication was which occurred in the process of solving the equation. If you come to such a principle of construction, all that you need plan and understand in detail is the primary automaton, and what you must furnish to it is a rather vaguely defined matrix of units; for instance, 1010 neurons which swim around in the cortex. But the thing becomes functioning only by being organized by the primary thing, and it is only the primary thing that you need plan. Then you begin to have this trait; you do not simply build-a primary machine to build a secondary one and then separate them. If you do not separate them but they stay in contact with each other, and if the functioning of the secondary machine presupposes that its contact is a primary one, then, I think that it is achievable that the thing can be watched by the primary automaton and be continuously reorganized when the need arises. I think that if the primary automaton functions in parallel, if it has various parts which may have to act simultaneously and independently on separate features, you may even get symptoms of conflict. Also, through that, since the secondary matrix is introducing things which are probably not rigorously defined in all details, the same primary automaton in two cases may not produce exactly the same structure. And, if the planning is well done, there will probably be a very high probability that the secondary thing will function the same way in all cases; but there will not be certainty, and, if you concentrate on marginal effects, you may observe the ensuing ambiguities. I just wanted you to realize that though these notions are vague, they indicate phenomena of this character. Especially when you go to much higher levels of complexity, it is not unreasonable to expect symptoms of this kind.