AN “EXTINCTION” PHENOMENON ON STIMULATION OF THE CEREBRAL CORTEX12
J.G. Dusser de Barenne and W.S. McCulloch
In the course of stimulation experiments on the motor cortex of the monkey an interesting phenomenon was met, which we wish to report.
The monkeys (macacus) were anesthetized with “Dial” and ether; in two experiments ether was used exclusively. The motor cortex was exposed for stimulation. The electrical stimulations, which were given for a few seconds, consisted of several patterns of various pulses, obtained from a thyratron stimulator after Schmitt,(1) 60 cycle alternating current or an ordinary inductorium. Both biphasic and half-rectified pulses were used. The durations of the various stimulatory periods and the intervals between them, though variable at will, were rigidly controlled by a mechanical device driven by synchronous A.C. motor. Unipolar and bipolar stimulation, polarizable and non-polarizable electrodes were used. The responses of the contralateral musculature were recorded with the isotonic method on a smoked paper kymograph. The anesthesia, the temperature of the animal and the external conditions of the cortex were kept as constant as possible over long periods of time. The wave form of the various stimulations was checked with the cathode ray oscillo-graph.
Under all of these experimental conditions the stimulations of a single focus of the motor cortex result in very constant responses, provided the intervals between the successive stimulatory periods are long enough. Then the only variation of the responses is that described in a previous paper(2) as “waves” or intrinsic fluctuations in cortical excitability. Usually, if very strong stimuli are avoided, intervals of half a minute to one minute are sufficient to obviate any disturbing influence resulting in variation of response, apart from the “waves” just mentioned. When the intervals between the stimulatory periods are reduced to 6 seconds or less the well-known phenomenon of primary facilitation, a marked increase in succeeding responses, promptly appears.
When the interval between two stimulation periods (stimulation frequency 53 per second) is taken at 13 seconds, leaving all other factors (shape, frequency and intensity of the pulses and duration of the periods of cortical stimulation) unchanged, the response to the second period is much smaller than that to the first, or it may be entirely absent. As soon as the interval between successive stimulations is augmented again to 27 seconds or more, the original size of the response promptly reappears. In other words, the response to a stimulation of the motor cortex, which alone would be fully effective, is small or absent if this stimulation is applied about 13 seconds after another equal stimulation. Some extinguishing factor
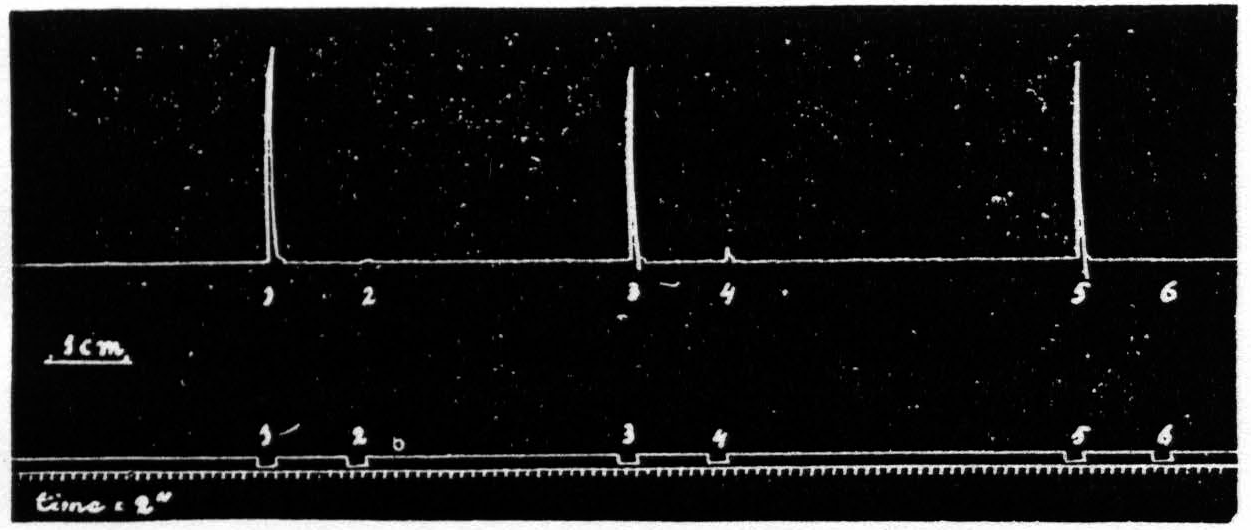
Figure 1: Exp. of Nov. 1, 1934. Monkey (macacus). “Dial” anesthesia. Three pairs of stimulation periods of one focus of left motor arm area, yielding extension of wrist: 1 and 2, 3 and 4, 5 and 6. Thyratron-tube discharges, resistance of 1000 ohms in series with animal. Duration, strength of stimulation, frequency and shape of pulses exactly the same in all 6 periods. Responses 2, 4, and 6 almost completely or completely extinguished because they are elicited 13 seconds after respectively antecedent stimulations. Stimulations 1, 3 and 5 fully effective, because of long preceding interval. Bottom line gives time in 2 seconds. Original size of graph may be determined by centimeter scale on left side.
must, therefore, be operative in the motor cortex or in the motor mechanisms involved, 13 seconds after an effective stimulation.
Whether this cortical extinction or inactivation which appears upon stimulation of a single motor focus is partial or total depends largely upon the strength of the stimulations. With strong stimuli the phenomenon does not occur or is minimal; with moderate stimuli it is obvious; with just supraliminal stimuli it is almost complete or total. It looks as if in the production of this extinction two antagonistic factors were effective: the one, excitation of the focus by the applied stimulus, the other, the extinguishing factor, active at about 13 seconds after the last antecedent stimulation. The time required for the recovery from the extinction set by the first cortical stimulation seems to be a function of the frequency of stimulation. For frequencies around 50 pulses per second it is found to be optimal at an interval of 13 seconds, for frequencies around 100, around 20 seconds.
This extinction phenomenon has been observed in all monkeys under observation—whether in deep or light anesthesia—since it was first noted, and at every focus tested of all the 3 major subdivisions (face, arm and leg) of the motor cortex (areas 4 and 6 of Brodmann).
The fact that extinction is greatest with slightly supraliminal stimulation is important, as it indicates that the phenomenon cannot be explained on the basis of fatigue or exhaustion of the cortex. If this were so, one would expect to find it more obvious with stronger stimuli and less evident with threshold stimuli. As has been stated. the reverse is true. The fact that decrease of the intervals between successive stimulations far within the duration of the optimal extinction interval, and, therefore, much closer to the refractory period, results in facilitation, is important, as it indicates that the phenomenon cannot be explained on the basis of a refractory period of the cortex following stimulation.
Extinction, as described here, is not to be confused with what is called cortical inhibition, which is cessation of a muscular response to stimulation of one focus of the motor cortex when a second, and antagonistic, cortical focus is concomitantly stimulated. Extinction occurs on stimulation of one motor focus following a preceding stimulation of the same focus, and that after the remarkably long interval of 13 to 20 seconds.
In conclusion it should be pointed out that this extinction is a physiological phenomenon, due to a functional change of the motor cortex under investigation or the motor mechanisms involved, and that it cannot be explained by physical inequalities of the various stimulations, because appreciable changes in polarization, resistance or impedance of the cortex are not responsible for the phenomenon. This has been shown by a new method of determining and recording of these various factors during electrical stimulation in living tissues, which will be published in the near future.
Footnotes
References
Schmitt, F.O., Science, 1932, 76, 328.
Brody, B.S., and Dusser de Barenne, J.G., Arch. Neur. and Psychiat., 1932, 28, 571.
For further research:
Wordcloud: Almost, Animal, Around, Completely, Conditions, Cortex, Cortical, Determined, Duration, Dusser, Effective, Electrical, Ether, Experiments, Explained, Extinction, Extinguishing, Factors, Focus, Following, Frequency, Function, Indicates, Interval, Mechanisms, Minute, Monkey, Motor, Optimal, Original, Paper, Per, Periods, Phenomenon, Pulses, Recorded, Resistance, Response, Result, Schmitt, Seconds, Shape, Stimulation, Stimulatory, Stimuli, Successive, Used, Various, Waves
Keywords: Cortex, Pulses, Periods, Phenomenon, Stimulations, Musculature, Animal, Time, Action, Ether
Google Books: http://asclinks.live/ybtq
Google Scholar: http://asclinks.live/9cf5
Jstor: http://asclinks.live/xszq