Options complexes:
Going beyond real options reasoning
Rita G. Mc Grath
Columbia University Graduate School of Business, USA
Max Boisot
INSEAD, ESP
Abstract
In business, as in nature, situational agility is a condition for access to resources and hence for adaptation and survival. But can strategy deliver such agility? And does the intentionality of individual players help to shape the strategy? Real options reasoning (ROR) theoretically secures a part for strategy to play, both in creating valuable resources and in adaptation. Yet because ROR adopts an essentially linear approach, it significantly understates both the value-creating and the adaptive potential of such a part. In this paper we argue firstly that those who draw upon ecological metaphors of organizations have largely ignored the role of strategic choice in value-creation and that ROR provides a good basis for the exercise of flexibility-preserving choices. Secondly we argue that the adroit use of multiple interacting options allows firms to harness the power of complexity thinking to the creation of value and to adapt to a greater range of environmental contingencies than is on offer in either the economic or financial treatment of options. Multiple interacting options thus extend the power of ROR to cover more complex and uncertain situations.
Introduction
The jury is still out on the question of strategic choice (Child, 1972). The strategy field takes it for granted that strategic choice influences outcomes, using as evidence the manifest heterogeneities in the performance of firms (Andrews, 1971; Selznick, 1957; Chandler, 1962; Ansoff, 1965). Others counter that such performance heterogeneity can be understood as reflecting differences in initial conditions coupled with fortuitous (or not so fortuitous) environmental conditions (Stinchcombe, 2000; Hannan & Freeman, 1989). If the latter focus on the external constraints of strategic choice, the former focus on how organizational actors might overcome them.
Central to this debate is the effectiveness of strategic foresight influences in shaping outcomes under what Cockburn, et al. (2000: 1128) call “a heavy cloud of uncertainty.” Scholars who question the value of such foresight point to limits on managerial rationality, to the tacit nature of organizational capabilities, and to the difficulty of changing organizational routines and processes (Levinthal & March, 1993; Hannan & Freeman, 1984; Christensen & Bower, 1996; Pfeffer & Salancik, 1978). The metaphors they use frequently recall natural processes such as biological evolution or embedded cognition, processes that occur without conscious intention or direction, and that exert overwhelming influence on choices and behaviors.
Scholars who favor the strategic choice perspective emphasize the sensitivity of managerial decisions to external information, the adaptiveness of many of their responses, and the differences in firm performance that result from different organization design choices (Galunic & Eisenhardt, 2001; Bower, 1970; Tushman & O’Reilly, 1997; Burgelman, 1996). Their metaphors are those of challenge and response (Toynbee, 1947), of navigating the rapids, of mastering capricious nature.
A more nuanced view of the role of strategic choice in value creation and adaptation can be developed by drawing on real options reasoning (ROR). An option is the right, but not the obligation, to take some specified action within some specified time frame. As a decision right, an option is at its most valuable under conditions of uncertainty when the future cannot be known. A real option applies financial options theory (with some modifications) to the case of real, i.e., non-financial, assets. All investments that extend in time yield their benefits in the form both of a flow of services and of either expanding or contracting decision rights. (Bowman & Hurry, 1993; Sanchez, 1993; Kogut & Kulatilaka, 1994a, 1994b). Decisions made later often benefit from more information and may thus be better―and hence more valuable―decisions, requiring less prior foresight. Finally, real options reasoning is a general way of thinking about resource allocation under conditions of uncertainty that exploits the provisions of option theory and its application to real options. That is, when allocating scarce resources, ROR develops specific mechanisms that safeguard the opportunities available for capturing potential upsides while affording some protection from the threat of downsides (Amram & Kulatilaka, 1999). In this sense, ROR both extends and must be distinguished from traditional risk management, which is usually exclusively concerned with downsides.
Because of its roots in the theory of financial options, ROR has been much concerned with the problem of how to value options. The resulting analytical bias as tended to limit the application of ROR to relatively simple and computationally tractable problems. In particular, it has confined ROR to a linear and additive treatment of the strategic choices that confront managers. In what follow, we shall ague that this seriously understates the potential that ROR offers entrepreneurial managers to exercise strategic choice, and that the adroit use of multiple interacting options can allow firms to create value and adapt to a wider range of environmental contingencies than would be the case if investments in firm-level capabilities were fixed and additive, as they are often portrayed in economic or financial treatment of options. In developing our argument, our aim will be to extend ROR in the direction of complexity theory, a new discipline that takes irreducible uncertainty as its key focus (Nicolis & Prigogine, 1989; Morin, 1973, 1980).
Like other organization and strategy scholars, we begin in section “The lesson from biology” by looking to biology for inspiration. Biology is another field in which scarce energetic and information resources have to be invested under conditions of uncertainty and change (Küppers, 1990; Brooks & Wiley, 1988). How does the process work there? We then introduce the idea of real options in section “Real options, choice and emergence” as an organizational analogue of choice-preserving systems in biology and show how, in the latter case, the efficacy of the processes result from the complexity of the interactions engaged in. In section “Constructing an options complex: An example” we discuss the implications of our analysis. A conclusion follows.
The lesson from biology
The evolutionary premises that underpin most critiques of the role of strategic choice are drawn from insights gained in the study of the adaptation and change of biological populations. Concepts originating in biology have had a significant impact on the study of organizations, most significantly in our understanding of how organizational populations form, develop and change (Hannan & Freeman, 1977; Boisot & Cohen, 2000). We look at two examples, one drawn from genetics and the other from immunology.
Genetics
The fundamental processes of evolutionary change are variation, selection, and retention (Aldrich, 1999; Campbell, 1965). In biology, variation is the product of genetic crossovers and random mutations occurring within interbreeding populations. A population evolves through natural selection, thus endowing the fittest organisms with a reproductive advantage (Mayr, 1978). Competition for scarce resources yields one selection process in which better-adapted organisms will triumph over less well-adapted ones within a given environment. Those of their characteristics that are ‘selected in’ are retained, or preserved in the population, and are then passed on to future generations. Biological selection finds its counterpart in the selection of business organizations in free markets. Businesses too create variations in terms of new products or services, and of structures and processes for delivering these. If a firm’s offerings fail to add value―thus allowing it to secure resources from its environment―it is ‘selected out’ in the face of competition.
Note that in biology, an organism may be short-lived, but still play an important evolutionary role, provided that it has survived for long enough to pass on its genetic endowment to the next generation. It is this genetic endowment rather than the individual organism that needs to be retained for evolutionary purposes. Biologists are thus careful to distinguish between the genotype of an organism and its phenotype. The organism’s genotype constitutes the total genetic endowment that it can potentially draw on, i.e., information available to the organism for its development―and pass on to succeeding generations. The organism’s phenotype then represents its observable characteristics, a physical implementation of its genotype in interaction with the different environments that it encounters as it develops. Thus, two animals with almost identical genetic endowments might still differ in size, shape, color, etc., reflecting differences in the environments which they were respectively exposed to.
In biological systems, selection does not operate directly on genotypes but on phenotypes (Mayr, 1978). If, for example, height conveyed a differential survival advantage, evolution favoring tall creatures would not occur unless the genes coding for height could be passed on from parent to offspring. The distribution of particular genotypes within a population would then, over time, come to reflect the differential advantage conferred up on the corresponding phenotypes by height.
Until the 1960s, evolution was treated as a purely additive phenomenon. Specific phenotypic traits were coded for by specific genes on a one-to-one basis. Evolution took place when, through mutation or genetic crossover―the product of mating―one gene replaced another, leading one trait to replace another. In this gene frequency view, the frequency of a given trait in a biological population would be matched by the frequency of the corresponding gene in the population’s genetic make-up (Fisher, 1930). Biologists, however, gradually came to hold a more elaborate view of the evolutionary process (Dobzhansky, 1962). In this view, genes are seen to work together in adaptive bundles or gene complexes. Genes do not necessarily map onto traits on a one-to-one basis and evolutionary change is no longer purely additive. Traits can arise from complex and flexible interactions between a limited number of genes to produce a wide variety of phenotypes (Kauffman, 1993). The combinatorial power of gene complexes thus generates a far greater amount of population-level diversity―and this, far more parsimoniously―than a one-to-one correspondence between a fixed allotment of genes and their expression in a population―the gene frequency view―might suggest.
Immunology
Edelman’s (1992: 75-78) studies of the immune system provides us with a second illustration from biology of how, with limited resources, combinatorial processes promote adaptation and generate diversity within a population. The job of the immune system is to combat bacteria and other foreign organisms that might represent a threat. It does this by triggering a highly specific two-part counterattack. It first synthesizes proteins called antibodies that combine with antigen molecules on the surface of the invader to destroy them. An antibody molecule binds the corresponding antigen like a key fitting a lock. Any individual has the capacity to produce a large but limited variety of antibody proteins that combine to match the set of ‘locks’ present on the surface of a foreign invader. When a foreign molecule enters the immune system, it encounters a population of antibody bearing cells (or lymphocytes) each with different antibodies on their surfaces. If the host organism can generate the right combination of antibodies, these will bind the invading molecule through its antigens and destroy it. In Figure 1, for example, a foreign body is bound to the appropriate lymphocyte cells (in this case 702, 735, and 886) and is destroyed by the combination of antibodies produced.
In the second part of the counterattack, the set of lymphocyte cells bearing the appropriate antibodies are then stimulated to divide repeatedly. Such ‘selecting in’ or ‘cloning’ of the relevant lymphocyte cells increases the population of antibodies which, when configured, have a good fit with the antigens of the invading molecule.
There are two major points to be made from this example. First, more than one type of lymphocyte is involved in fending off the invader―they work in teams. So in Figure 1, instead of there being only 11 possible responses, one for each lymphocyte, there is actually a vast number of combinatorial responses that the immune system can summon from a limited base. Second, instead of the organism being at the mercy of whatever immunological resources it was born with, it is itself able to develop and extend these resources, and hence its combinatorial capabilities. An individual organism’s immune system in effect evolves to combat the specific contingencies it faces as it develops and grows. Consequently, no two organisms have identical sets of antibodies. Figure 2 illustrates the point.
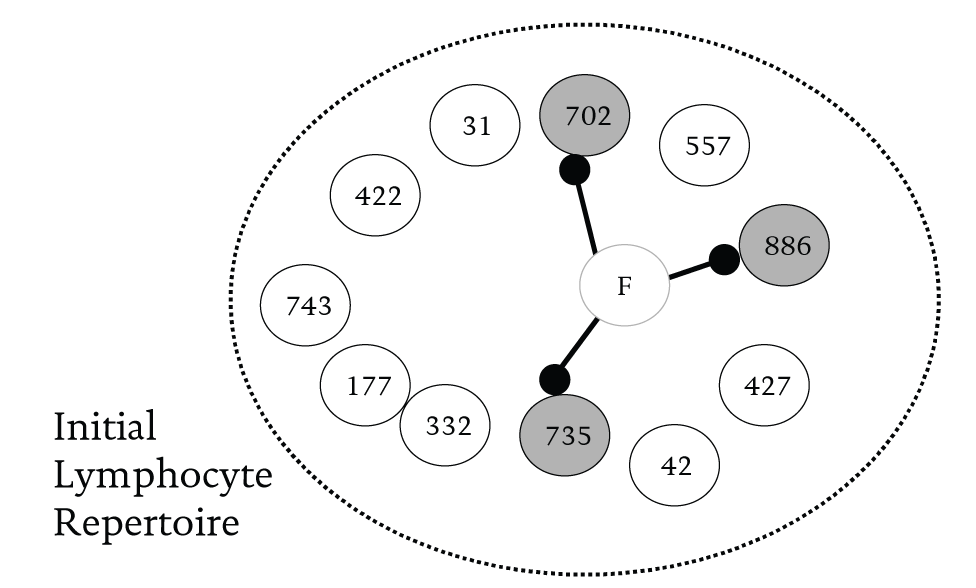
Figure 1 Foreign molecule F bound to antibodies on lymphocytes that fit parts of its shape (Small black dots represent antigens with a binding site that fits the antibody surface)
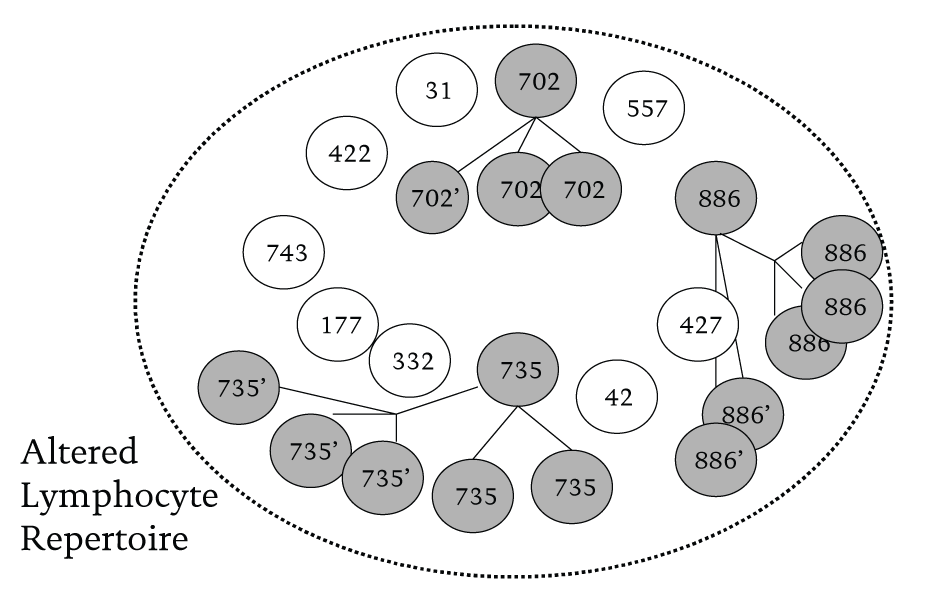
Figure 2 Affected lymphocytes (702, 886 and 735) are stimulated to make clones and increase their numbers in the population of cells in an individual’s immune system. These clones also all have the potential to mutate to say 702’,735’,886’.
Biology and complexity
The cybernetician, William Ross Ashby, argued that, if it is to avoid disintegration, the variety mobilized by any system, living or otherwise, must match that of the environment it finds itself in, i.e., its internal complexity must be at least equal to that of the external complexity that confronts it. This argument became known as Ashby’s law (Ashby, 1956). In the gene-frequency model, requisite variety demanded a substantial genetic repertoire: each conceivable environmental contingency required its own matching gene. In the gene-complex model, by contrast, variety is achieved by leveraging the combinatorial power of the genes in the genotype, thus increasing the adaptive capacity of a limite d repertoire of gene s exponentially. The current thinking in evolutionary biology is that the gene-frequency and the gene-complex models actually operate in tandem in achieving requisite variety. Evidence is accumulating that because of the combinatorial capability of the genotype, the genome is actually capable of dealing with far more variety than had originally been supposed. The same argument applies in the case of the immune system. A limited number of antibodies combine in myriad ways and multiply to ward off a much larger variety of antigens.
In sum, what do gene complexes and antibodies have in common? Both leverage complex interaction between a limited number of elements―genes, antibodies―to mobilize a much wider range of innovative or adaptive responses than a purely additive view of such elements would lead one to expect. The complexity of these interactions arises from the degrees of freedom―the flexibility―available to these elements. We can think of them as having options to combine that will be exercised under certain envi-ronmental contingencies. In both the genetic and the immune system we see a biological process leveraging limited resources in nonlinear yet parsimonious ways, i.e., harnessing complexity (Axelrod & Cohen, 1999).
Biological analogies and organizational strategies
In translating ideas from biological populations to organizational ones, scholars in the organization ecology movement have specified a number of boundary conditions (Baum, 1996). First, the genetic endowment of a firm―taken here as its routines and competencies―is inseparable from the organizational whole, so that organizational genotypes and phenotypes cannot be clearly distinguished. Second, the amount of variety that a given organization can adapt to as it develops is more or less fixed by the initial endowments and choices of its early years. In unforgiving environments, therefore, firms cannot change either their endowments or their strategies fast enough to survive. Not only are a firm’s resources construed as being fixed by initial endowment, they are also construed as being relatively inflexible. No combinatorial potential is on offer to expand the resource base of the initial endowment. The firm’s response to environmental change is thus taken to be strictly linear and additive, with little or no scope for strategic choice or for organizational learning. Third, for this reason, organizational fitness is usually modeled as random. As Hannan and Freeman (1984: 150) put it “in a world of high uncertainty, adaptive efforts ... turn out to be essentially random with respect to future value.” In the organizational ecology perspective, then, change is at best disruptive and at worst highly dangerous.
Some scholars have challenged this rather narrow application of biological analogies to firms, arguing that internal processes of variety generation and selection (such as design and planning) can create a potential for adaptation beyond what might be secured by random variation and selection pressures alone (Meyer, 1990; Eisenhardt & Tabrizi, 1995; Levinthal, 1997). They do not claim that the resulting outcomes are necessarily intentional; they may well be emergent (Mintzberg & Waters, 1982). Yet to ignore them is as one-sided as to assume that successful organizational adaptation is exclusively the product of managerial foresight.
In effect, in applying evolutionary analogies to the study of organizations, scholars have overlooked two ideas that are central to an understanding of many biological processes. Firstly, interactive complexes of genes, lymphocytes or other biological entities offer a potential for adaptation and innovation that goes significantly beyond what can be achieved by a linear combination of the same entities. For organizations, the internal coordination processes that link together rules and routines, often under strategic direction, could be designed to build up such interactive complexes. Think, for example, of how a multidisciplinary project team might quickly be pulled together from different parts of a consulting organization and configured so as to respond speedily and flexibly to an unforeseen opportunity or threat.
The second idea is that selection processes do not operate on individual biological entities alone, but on complexes of these entities. When we apply this idea to routines and competencies, the implication is that inferior routines might well survive if the complex of which they are a part is not selected out while superior routines might be selected out if the complex with which they are associated fails the selection process. This suggests two roles for strategic choice in the organization. The first is as shaper of routines and competencies into adaptive complexes. It is the capacity to exploit complex interactions between elements of the resource base that allows it to expand in emergent ways. The second is as custodian for the selection processes that will favor the development of certain adaptive processes over others (Burgelman, 1996). Pragmatically, this has to do with resource allocations within the firm. The lesson from biology is that, viewed through the complexity lens, such an allocation can build a much broader and flexible repertoire of responses from a limited resource base than was assumed possible.
In short, the strategic choices made by managers―whether consciously or unconsciously exercised―can influence, even if it cannot determine, how a firm’s rules and routines will be created and how they will be selected. As we shall argue next, such choices are often in effect creating complexes of interacting options. If managers can be made aware that this is what they are doing, they will be able to do it better.
Real options, choice, and emergence
Bowman and Hurry (1993) were influential in suggesting that the incremental choice process so evident in the emergence of organizational strategy could be viewed through an options lens. They argued that the resources of a firm contain within them latent potential to create strategic value, which they termed shadow options. Depending upon the results of the options exercise, a firm might continue its strategy, change strategic direction by terminating its options or allowing them to expire or invest in new options elsewhere. The process of creating, recognizing and exercising options that they depict bears a strong resemblance to the processes of gene expression and of clonal selection in biology. As in the biological case, a manager must recognize the opportunity to exercise options. That is, in the process of organizational evolution and adaptation, he or she acts as an agent of selection.
Nevertheless Bowman and Hurry promote a strictly additive view of option. By arguing that options, like genes and like antibodies can be configured and made to work as adaptive complexes, we go beyond this additive perspective. Indeed, drawing on our two biological examples, we claim that networks of options interacting in complex ways , i.e., as options complexes, will display nonlinear emergent properties that are potentially far more value adding than the readily identifiable synergies achievable through creating and exercising Bowan and Hurry’s shadow options. Indeed, the size of the upsides achievable and of the downsides forestalled through the emergent properties of options complexes make a concern with option valuation quite secondary (see below). In contrast to risk management, the options complexes approach exploits the opportunities latent in high degrees of uncertainty instead of attempting to reduce that uncertainty. Nevertheless, our augmented version of ROR―which we shall label AROR―acts as a complement to risk management rather than as a substitute for it.
AROR allows organizations to operate at a higher level of uncertainty than they might otherwise be able to handle, even under ROR. Options complexes, like the gene complexes or teams of lymphocytes in the immune system, can activate a much wider variety of choices, from the limited number of rules and routines that reside in an organization’s repertoire, than is suggested by conventional ROR. Properly configured, they thus offer the potential to both create unexpected new forms of value, as well as to significantly increase the range of environmental variations that can be addressed. Uncertainty now becomes a source of creative emergence that escapes all attempts at valuation―one of the main preoccupations of ROR. Indeed, given its particular analytical focus, what has been missing from ROR is any sense of the creative potential of strategic choice.
New value creation in an options framework
We have so far not addressed a major issue in real options theory, namely the relationship between the presence of options and the value of a firm. Myers, (1977) and Myers and Turnbull (1977) were among the first scholars in the field of finance to argue that part of the value of a firm derives from intangible assets or “options to purchase additional units of productive capacity in future periods” (Myers & Turnbull, 1977: 331-332). The value of a firm, in other words, cannot simply be understood by assessing the present value of the future cash flows it is likely to generate from ongoing operations. Instead, some portion of firm value stems from as-yet unknown future opportunities. That portion of value stems directly from the presence of options. The total value of a firm at any given point in time can thus be understood as a mix of resources that are amenable to present-value calculations, plus those resources that investors credit with the potential for creating new value in the future, or option value. This ROR perspective on a firm’s value is shown in a simplified form in Figure 3.
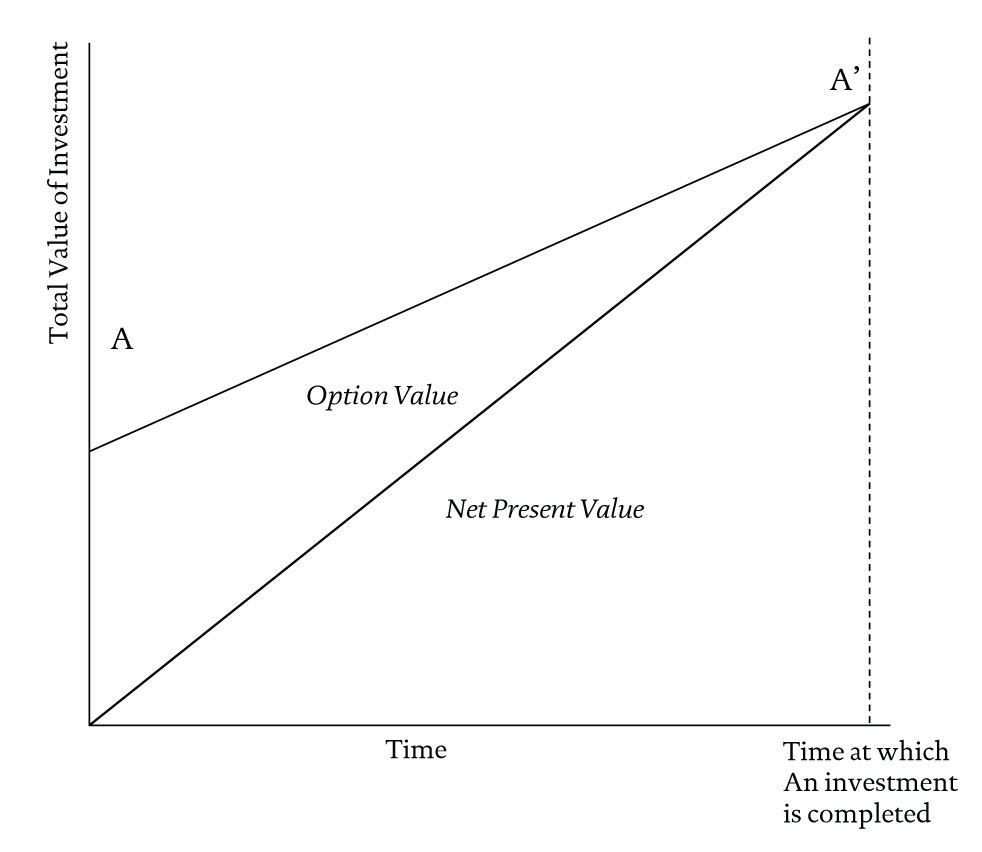
Figure 3 ROR at Work
In the initial founding stages of a firm, virtually all the value that it represents consists of option value, because those combinations of assets and routines that might generate predictable future cash flows have not yet been created. The founding and organizing process of a firm result in the founding team making choices and investments such that the proportion of present value increases. To the extent that these choices create greater value than their pre-investment worth, a founder has successfully created what Rumelt, (1987) calls “entrepreneurial rents,” which are the returns generated by taking action in the face of uncertainty (see also Knight, 1921)1.
The vertical axis of the figure represents the total value to be had from any given investment. The curve AA’ represents different mixes of present value and option value in the total value of a firm. On the left-hand side of the diagram we meet the pure entrepreneur. The pure entrepreneur is all option value and no present value to speak of, with no investible assets around which to construct a present value calculation. The presence of managerial (in this case founding) discretion can be detected in the ways in which entrepreneurs create real assets by judiciously exercising options, by moving towards the right in the diagram and by creating net present value where none previously existed. The move to the right represents the steps in an investment project in which, through the successive exercise of options, option value gradually gives way to net present value.
Note that the investment process just described is a purely linear one. The options that are exercised are sequentially arranged and additive in their effects. We are still in the world of conventional ROR. How might AROR change the picture?
A key skill for entrepreneurs consists of the ability to construct deals. When making deals, heterogeneous bundles of resources are pulled together and configured in such a way that the resulting whole becomes worth more than the sum of the parts. Those engaged in deal-making seek to capture the upside potential of nonlinear outcomes, while avoiding commitment to downside risk. The resources required are not necessarily owned by the deal-maker. Indeed, a popular definition of the entrepreneur is an individual that seeks opportunity without regard to resources currently possessed (Stevenson & Jarillo, 1990).
The essential skill consists of weaving conditional value-adding connections between different types of resources so as to favor emergent outcomes. In effect, a deal-making entrepreneur is creating the equivalent of the genetic endowment of the organization, by building up a network of options that can be activated under certain circumstances. The underlying asset being created is itself often nothing more than the network of interrelated options, i.e., an options complex, that the deal-maker is putting together. Such a network of interrelated options creates upside potential when the connections are activated and, much like a diversified insurance portfolio, defrays risk when they are not. This is the AROR perspective. To the value established by ROR it adds an emergent value created by complex nonlinear interactions between options. This is illustrated in Figure 4.
Constructing an options complex: An example
We illustrate AROR at work with the following toy example. To add force to our argument, we take the case of a pure entrepreneur, i.e., someone with imagination but few other initial resources. Imagine, then, an aspiring real-estate developer with limited resources living in a growing city. She observes that changes are about to take place in the city’s central business district. A new rapid transit system is under discussion but the city authorities have not yet decided where to locate the station stops. One stop under discussion might be within a residential area. If it is built, then the zoning laws are likely to change to permit more land to be placed in commercial use. Commercial activity has been on the increase, while the residential population has been declining―indeed within one block, two applications have just been put in to convert existing houses to commercial use. The outcome of the planning applications on the two existing houses can serve as a bellwether for where the area is heading. If permission is granted, other investments are likely to follow. How might the aspiring-yet-resourceless developer take advantage of such an uncertain opportunity?
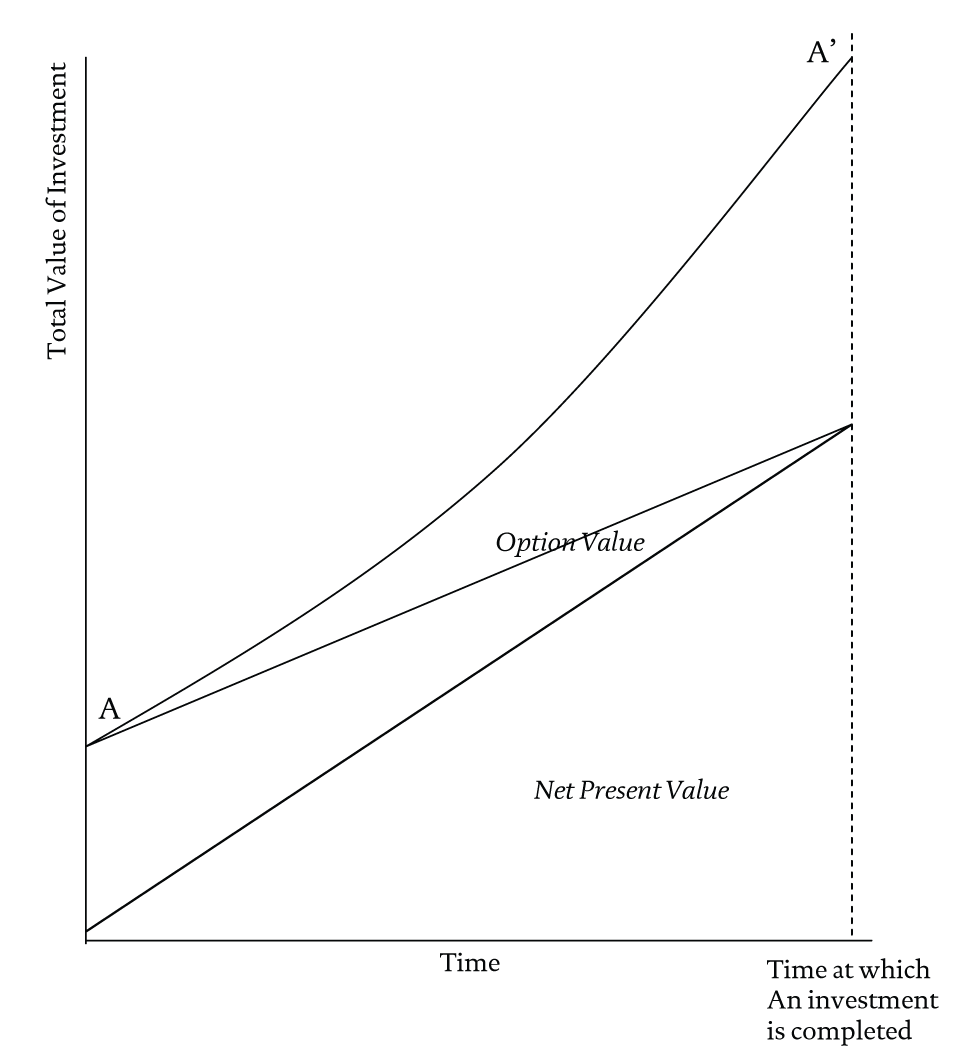
Figure 4 AROR at Work
Imagine the following sequence of deals:
- The entrepreneur negotiates with owners of the two properties under planning application to take out an option to buy them, contingent upon their being granted planning approval. As an incentive, the entrepreneur might offer a price premium over the market price of the properties in their new use.
- Next, she negotiates with other residential property owners for options to buy their properties, also for a premium. If planning permission is granted on the original two properties, a zoning precedent has been set for the block as a whole. Development of the block as a whole would not only be worth many times more than the sum of piecemeal developments, it would also act as a trigger for the renewal process of the area in general.
- She then negotiates with city authorities to place a mass transit stop adjacent to the relevant block, contingent on a certain volume of commercial development.
- Next, the entrepreneur takes out options to buy out residents in neighboring blocks, options which she will only exercise after a decision has been made on the location of the rapid transit station. This will allow her to capture the improved land values created by the development of one block and to use it as collateral.
- She now obtains a conditional offer of project funding from a bank secured against the improved land values created by any planning approvals.
- Finally, she takes the whole options complex so far created to an established developer and negotiates for funding for the purchase of the options in return for a substantial minority share of the equity in the development.
In such a network of deals, the entrepreneurial developer invests time but not money. Yet by the end of step 5 she has created a marketable asset. Her time investment has secured a nonlinear payoff, even if she does not carry out the development herself. And the process of creating, configuring, and exercising options can be repeated once the project is underway―exploiting developments in the vicinity that have been triggered by her initiative, looking at adjacent rapid transit stations for spillover opportunities, etc.―to keep the option value of the project at some optimum level relative to its net present value (NPV).
In the field of entrepreneurship, construction of deals that reflect parsimonious investments to contain downside risk and to access significant upsides are legion and well documented (Hambrick & MacMillan, 1984; Starr & MacMillan, 1990). Microsoft Chairman Bill Gates, for instance, risked only $50,000 to acquire the technology that became MSDOS, the platform for Microsoft’s rocketing growth after another deal was struck with IBM to use it as the operating system for its personal computers (Cringely, 1996).
The move from left to right in the diagram as an investment project progresses in Figure 3 has the effect of decreasing the ratio of potential choices a decision maker has (constituting option value) to commitments that have been made (amenable to present value analysis). Over time, as options are exercised, the network of options becomes increasingly fixed. Live options that remain come to constitute small islands of discrete individual choices in a sea of commitments. In effect, we revert back to the gene- frequency model as we move toward the right in the diagram, and the move itself is experienced as an increase in the number of constraints on one’s ability to choose. On the far right of the diagram, choice has effectively been expunged from the system.
The gradual loss of flexibility entailed by a move to the right in the diagram reduces an organization’s ability to learn and adapt. Inertia builds up. The further to the right an organization moves in the diagram, therefore, the greater the risk that it will be selected out by changes in its environment. Thus our account so far is fully compatible with the one advanced by organizational ecology as well as with conventional ROR thinking. Figure 4, however, tells a different story. Whether or not option value is eroded as one progresses through an investment project depends on how the project is construed. Through a skillful and continued creation and configuration of options one can effectively maintain or expand the option value of the project as an investment with little or no sacrifice of NPV.
The message of Figure 4 is that there is nothing inevitable about the progressive fossilization of choice implied by Figure 3. While older organizations are more likely to find themselves trapped in the ROR logic of Figure 3―after all, they will have accumulated assets―they retain the potential to switch to an AROR logic. We associate this logic with a willingness to engage in exploratory learning. Conventional ROR logic is more tilted towards exploitative learning (March, 1991).
Implications
Both genetics and immunology leverage complex interactions between a limited number of elements―genes, lymphocytes―to deliver a degree of flexibility, adaptation and innovation that far exceeds what economic organizations operating in the human sphere are capable of, using a conventional ROR approach. This approach, the one that currently predominates in both finance and strategy, is analogous to the gene frequency approach. Here, options tend to be viewed as discrete, linear, and additive. In effect, they are treated as independent members of a portfolio to which they can be added or subtracted with little or no effect on the other members of the portfolio. Each specific contingency is dealt with by taking out one option designed to cover an uncertain but highly circumscribed context. Any given option is then either exercised or allowed to lapse when the relevant uncertainties have been resolved with little or no effect on other options in the portfolio. Two assumptions underpin the approach. Firstly, that the assets and the contingencies covered by the option are clearly identifiable, and secondly that the signals that would trigger its exercise or an exit are clear. Under these conditions a gene frequency approach makes sense.
However, it is not far removed from what risk management or decision theory would advocate. If managers are required to focus exclusively on identifiable possibilities, ones to which a probability distribution might be attached―even if it is a flat one―it is hardly surprising that the number of outcomes that can be managed is severely circumscribed. Yet where the full range of possible outcomes cannot be so readily identified or circumscribed, investing in a sufficient number of options to cover all possible contingencies could rapidly lead an organization to exhaust its resources. In highly complex or turbulent environments, a mechanism for improving the ratio of contingencies covered to options becomes essential and for this, the options-complex approach suggested by AROR has more to offer. In complex and turbulent environments, where analysis and prediction lose their purchase, options are likely to be at their most effective in confronting uncertainty when configured into meaningful combinations, i.e., they benefit from ‘teamwork’. The entrepreneurial manager leverages the combinatorial potential of a firm’s options-complex.
Managerial discretion is manifest in investment choices that position the resources in a firm across the spectrum of value represented in Figure 3. In both finance and in population ecology, one might expect managers to invest in a portfolio of options according to the precepts of ROR indicated in the figure. Here managerial choice is front-loaded and flexibility declines as options are exercised, to deliver the structural inertia predicted by organizational ecologists. Conventional finance places a premium on present value that has the effect of discounting the structural inertia identified by organizational ecologists. Inertia is for the future! In neither field are the roles of strategic choice and flexibility deemed to be particularly central. The AROR curve shown in Figure 4 represents the new insights and possibilities brought about by complexity thinking. Here, flexibility and option value is maintained throughout and, indeed, can be made to increase. If, one could clearly articulate and legitimate AROR, a form of reasoning that many entrepreneurial managers intuitively or unconsciously subscribe to and act upon, then strategically minded investors would feel more comfortable investing in the creation of resources that offer a higher proportion of option value to NPV than ROR can deliver. In other words, investors would themselves come to adopt a more entrepreneurial orientation, one that places the strategic choices made by managers centre-stage.
To be sure, there is mounting empirical evidence that strategic investors respond to the presence of option value. Kogut, (1991) observed that investment behavior in joint ventures was sensitive to signals of change in future value of the venture, responding to signals to (in effect) exercise options to expand and acquire. Dixit, (1992) studied hysteresis effects, observing that managers stay in poor positions longer and insist on higher hurdle rates for investment than a strict present value analysis would suggest they should. McGrath and Nerkar (2001) found that pharmaceutical firms invested in patenting in a manner that was consistent with a desire to balance option value and present value, for instance slowing their exploration of new fields when older ones showed promise.
But this is all evidence of ROR. Our point is that ROR, as currently formulated, will not help managers meet the challenges of an ever more complex world―those of ever more rapid technological change, ever increasing globalization, ever intensifying competition, ever shorter product life cycles, etc. To address the problem, in this paper we add to ROR a complexity dimension, arguing that the theory of real options, augmented by complexity thinking, can help scholars understand how environmental constraint and strategic choice intersect. Just as advocates of extremely limited managerial discretion often do, we have drawn from theories of adaptation applied to biological evolution. We have shown, however, that in certain fields in biology, such as genetics and immunology―the list is growing―complexity thinking offers far more innovative and adaptive potential than the more linear forms of thinking associated with conventional ROR can deliver. By analogy, we suggest that networks of interacting options can help entrepreneurs and managers both create new value and adapt value-generating strategies to a wider range of contingencies than is typically envisaged in the literature.
Although ROR has been applied primarily to strategic investment decisions, it turns out that such reasoning constitutes a dimension of all decision-making that is characterized by irreversibilities, uncertainties, and flexibilities in the timing of decisions (Dixit & Pindyck, 1994). Every decision that a manager or entrepreneur takes which has these characteristics has an option value as well as a net present value. An options approach can thus affect how one thinks about many if not most things that one is called upon to decide on as a strategist. Even in the absence of the ability to predict future outcomes, intelligence can still be applied to the question of value creation for the long run through the construction of contingent options. But as the world becomes more complex―through the effects of greater connectivity and the globalization that results―we will discover that the flexibility on offer through conventional ROR will not allow us to rise to the challenges. Firms need to match external complexity with internal complexity without blowing their budgets and depleting themselves of resources. AROR, by combining ROR with complexity thinking, suggests how the challenge might be approached.
The entrepreneurial ability to flexibly configure investment opportunities into option complexes constitutes an asset in its own right―albeit an intangible one. Like other types of intangible assets, entrepreneurial and managerial capabilities of this kind can neither be directly traded nor tracked in capital markets. A fascinating opportunity for future research lies in developing better ways of assessing the value of such intangible assets and capabilities. We suspect that the answer will require a better exploitation of the new theories of complexity.
In sum, we have sought to argue for a more nuanced theory of strategic choice in value creation and adaptation than is conventionally used either in organizational ecology or in strategy. Ecological metaphors have largely ignored intentionality in entrepreneurial value-creation, leaving unexplained entire classes of phenomena. How, for instance, can the ecological literature explain the persistence of the so-called serial or habitual entrepreneur? Absent some notion of intentionality in the creation of value, it is hard to understand how such people can succeed in multiple ventures, often located in different product/market spaces (Starr & Bygrave, 1992; Wright, et al., 1999). Both ROR and AROR provide an economic rationale for investing in uncertain exploration. Both offer reasonable mechanisms to achieve a balance between exploration and exploitation advocated by March, (1991). AROR, however, by departing from conventional treatments of options as discrete, additive and linear, and by embracing complexity thinking, shifts the balance more forcefully towards the exploration end of the spectrum.
Conclusion
ROR became popular in the field of strategy after Fisher Black and Myron Scholes had shown how to value financial options. Much of ROR’s focus has been on option valuation. The computational requirements imposed by this concern with valuation has forced a more discrete treatment of real options than was necessary for using them. In this paper we have been concerned with a particular way of thinking about options, not a particular way of valuing them. We believe that once the combinatorial power of options is acknowledged, appropriate computational strategies for valuing them will follow. These are unlikely ever to achieve the simple mathematical elegance of the Black-Scholes equations that are used to evaluate financial options. They should nevertheless offer decision makers useful ways of going beyond what remain at present merely hazy intuitions concerning the need for flexibility when making investments. Drawing on biological thinking can help us to realize that AROR offers important tools of organizational adaptation to an increasingly uncertain and rapidly changing world, tools that help to deal with external complexity with limited means, tools that have their counterpart in natural systems facing similar challenges.
Note
References
Aldrich, H. (1999). Organizations evolving, Thousand Oaks, CA: Sage Publications, ISBN 0803989199.
Amram, M. and Kulatilaka, N. (1999). Real options: Managing strategic investments in an uncertain world, Boston, MA: Harvard Business School Press, ISBN 0875848451.
Andrews, K. R. (1971). The concept of corporate strategy, Homewood, IL: Irwin, ISBN 0256183295 (1994).
Ansoff, H. I. (1965). Corporate strategy, New York, NY: McGraw Hill, ISBN 0283993545 (1986).
Ashby, W. R. (1956). An introduction to cybernetics, London, UK: Chapman and Hall, Ltd., ISBN 0416683002 (1964).
Axelrod, R. and Cohen, M. (1999). Harnessing complexity: Organizational implications of a scientific frontier, New York, NY: The Free Press, ISBN 0465005500 (2001).
Baum, J. A. C. (1996). “Organizational Ecology,” S. R. Clegg, C. Hardy, W. R. Nord (eds.), Handbook of organization studies, ISBN 0761951326, Thousand Oaks, CA: Sage Publications, pp. 77-114.
Boisot, M. and Cohen, J. (2000). “Shall I compare thee to...an organization?” Emergence, 2(4): 113-135.
Bower, J. L. (1970). Managing the resource allocation process: A study of corporate planning and investment, Boston, MA: Harvard Business School Press, ISBN 0875841287 (1986).
Bowman, E. H. and Hurry, D. (1993). “Strategy through the option lens: An integrated view of resource investments and the incremental-choice process,” Academy of Management Review, ISSN 0363-7425, 18: 760-782.
Brooks, D. R. and Wiley, E. O. (1988). Evolution as entropy: Towards a unified theory of biology, Chicago: University of Chicago Press, ISBN 0226075745.
Burgelman, R. (1996). “A process model of strategic business exit: Implications for an evolutionary perspective on strategy,” Strategic Management Journal, ISSN 10970266, 17: 193-214.
Campbell, D. T. (1965). “Variation and selective retention in socio-cultural evolution,” in H. R. Barringer, G. I. Blanksten and R. Mack (eds.), Social change in developing areas: A reinterpretation of evolutionary theory, Cambridge, MA: Schenkman, pp. 19-49.
Chandler, A. (1962). Strategy and structure: Chapters in the history of the industrial enterprise, Cambridge, MA: MIT Press, ISBN 0262530090.
Child, J. (1972). “Organizational structure, environment
and performance: The role of strategic choice,” Sociology, ISSN 0038-0385, 6: 1-22.
Christensen, C. and J. Bower. (1996). “Customer power, strategic investment, and the failure of leading firms,” Strategic Management Journal, ISSN 1097-0266, 17: 197-219.
Cockburn, I., Henderson, R. and Stern, S. (2000). “Untangling the origins of competitive advantage,” Strategic Management Journal, ISSN 1097-0266, 21: 1123-1145.
Cringely, R. X. (1996). Accidental empires: How the boys of Silicon Valley make their millions, battle foreign competition and still can’t get a date, New York, NY: Harper Collins, ISBN 0887308554.
Dixit, A. K. (1992). “Investment and hysteresis,” Journal of Economic Perspectives, ISSN 0895-3309, 6: 107-132.
Dixit, A. K. and Pindyck, R. S. (1994). Investment under uncertainty, Princeton, NJ: Princeton University Press, ISBN 0691034109.
Dobzhansky, T. (1962). Mankind evolving: The evolution of the human species, New Haven: Yale University Press, ISBN 0300004273.
Edelman, G. (1992). Brilliant air, brilliant fire, Middlesex, UK: Penguin Books, ISBN 0465007643 (1993).
Eisenhardt, K. and Tabrizi, B. (1995). “Accelerating adaptive processes: Product innovation in the global computer industry,” Administrative Science Quarterly, ISSN 00018392, 40: 84-110.
Fisher, R. (1930). The genetical theory of natural selection, Oxford, UK: Oxford University Press, ISBN 0198504403 (2000).
Galunic, D. C. and Eisenhardt, K. L. (2001). “Architectural innovation and modular organizational forms,” Academy of ManagementJournal, ISSN 0001-4273, 44(6): 12271247.
Hambrick, D. C. and MacMillan, I. C. (1984). “Asset parsimony: Managing assets to manage profits,” Sloan Management Review, ISSN 0019-848X, 25 (Winter): 67-74.
Hannan, M. and Freeman, J. (1989). Organizational ecology, Cambridge, MA: Harvard University Press, ISBN 0674643488.
Hannan, M. and Freeman, J. (1984). “Structural inertia and organizational change,” American Sociological Review, ISSN 0003-1224, 49: 149-164.
Hannan, M. T. and Freeman, J. (1977). “The population ecology of organizations,” American Journal of Sociology, ISSN 0002-9602, 82: 929-964.
Kauffman, S. (1993). The origins of order: Self-Organization and selection in evolution, Oxford, UK: Oxford University Press, ISBN 0195079515.
Knight, F. H. (1921). Risk, uncertainty andprofit, Chicago, University of Chicago Press, ISBN 1587981262.
Kogut, B. and Kulatilaka, N. (1994a). “Operating flexibility, global manufacturing, and the option value of a multinational network,” Management Science, ISSN 0025-1909, 40: 123-139.
Kogut, B. and Kulatilaka, N. (1994b). Options thinking and platform investments: Investing in opportunity,” California Management Review, ISSN 0008-1256, 36: 52-71.
Kogut, B. (1991). “Joint ventures and the option to expand and acquire,” Management Science, ISSN 0025-1909, 37: 19-33.
Küppers, B.-O. (1990). Information and the origin of life, Cambridge, MA: MIT Press, ISBN 026211142X.
Levinthal, D. (1997). “Adaptation on rugged landscapes,” Management Science, ISSN 0025-1909, 43: 934-950.
Levinthal, D. and March, J. G. (1993). “The myopia of learning,” Strategic Management Journal, ISSN 01432095, 14: 95-112.
March, J. G. (1991). “Exploration and exploitation in organizational learning,” Organization Science, ISSN 1047-7039, 2: 71-87.
Mayr, E. (1978). “Evolution,” Scientific American, ISSN 0036-8733, 239: 46-55.
McGrath, R. G. and Nerkar, A. (2001). “Real options reasoning and a new look at the R&D investment strategies of pharmaceutical firms,” Strategic Management Journal, ISSN 0143-2095, 25(1): 1-21.
Meyer, M. (1990). “Notes of a skeptic: From organizational ecology to organizational evolution,” in J. Singh (ed.), Organizational evolution: New directions, Newbury Park, CA: Sage Publications, ISBN 0803936591, pp. 298-314.
Mintzberg, H. and Waters, J. A. (1982). “Tracking strategy in an entrepreneurial firm,” Academy of Management Journal, ISSN 0001 4273, 25: 465-499.
Morin, E. (1973). Le paradigme perdu: La nature humaine, Paris: Editions du Seuil, ISBN 2020053438 (1979).
Morin, E. (1980). La methode: 2, La Vie de la Vie, Paris: Editions du Seuil, ISBN 2020056372.
Myers, S. C. (1977). “Determinants of corporate borrowing,” Journal of Financial Economics, ISSN 0304-405X, November: 147-176.
Myers, S. C. and Turnbull, S. M. (1977). “Capital budgeting and the capital asset pricing model: Good news and bad news,” The Journal of Finance, ISSN 1540-6261, 32: 321-333.
Nicolis, G. and Prigogine, I. (1989). Exploring complexity: An introduction, New York, NY: W. H. Freeman and Company, ISBN 0716718596.
Pfeffer J. and Salancik, G. (1978). The external control of organizations: A resource dependence perspective, New York, NY: Harper & Row Publishers, ISBN 080474789X (2003).
Rumelt, R. P. (1987). “Theory, strategy and entrepreneurship,” in D. J. Teece (ed.), The competitive challenge: Strategies for industrial innovation and renewal, New York, NY: Harper and Row, ISBN 0887304028 (1990), pp: 137-158.
Sanchez, R. (1993). “Strategic flexibility, firm organization and managerial work in dynamic markets: A strategic options perspective,” in P. Shrivastava, A. Huff and J. Dutton (eds.), Advances in Strategic Management, Volume 9, Greenwich, CT : JAI Press, ISBN 1559386479, pp. 251-291.
Selznick, P. (1957). Leadership in administration: A sociological interpretation, New York, NY: Harper and Row.
Starr, J. A. and MacMillan, I. C. (1990). “Resource cooptation and social contracting: Resource acquisition strategies for new ventures, Strategic Management Journal, ISSN 0143-2095,11:79-92.
Starr, J. and Bygrave, W. D. (1992). “The second time around: Assets and liabilities of prior start-up experience,” in S. Birley, I. C. MacMillan and S. Subramony (eds.), International perspectives on entrepreneurship research, North-Holland, ISBN 0444895264, pp. 340-363.
Stevenson, H. H. and Jarillo, J. C. (1990). “A paradigm of entrepreneurship: Entrepreneurial management,” Strategic Management Journal, ISSN 0143-2095, 11: 17-27.
Stinchcombe, A. L. (2000). “Social structure and organizations: A comment,” in J. Baum and F. Dobbin (eds.), Economics meets sociology in strategic management: Advances in strategic management, Greenwich, CT: JAI Press, ISBN 0762306645, pp. 229-259.
Toynbee, A. (1947). A study of history, abridgement by D. C. Somerville, Oxford, UK: Oxford University Press, ISBN 0195050800 (1987).
Tushman, M. L. and O’Reilly, III, C. A. (1997). Winning through innovation: A practical guide to leading organizational change and renewal, Boston, MA: Harvard Business School Press, ISBN 1578518210 (2002).
Wright, M., Westhead, P. and Sohl, J. (1999). “Editor’s introduction: Habitual entrepreneurs and angel investors,” Entrepreneurship, Theory and Practice, ISSN 1042-2587, 22: 5-21.