Complexity:
A Maturanian Perspective
Peter Bond
University of Liverpool, ENG
Abstract
Biologists and cyberneticians Humberto Maturana and Francisco Varela over a period of some 30 years developed a paradigm challenging body of work known as autopoietic theory or the biology of cognition, which Fritjof Capra wrote of as one of three foundational pillars of complexity science. In practice this position was never attained, and wasn’t widely recognized. The objective of this article is to attempt to reintegrate Maturana and Varela with complexity science by demonstrating its relevance to current thinking from the emerging field of philosophy of complexity led by the late Paul Cilliers.
Introduction
Fritjof Capra developed his vision of a new paradigm of complexity science in a series of books, Turning Point (1990), The Web of Life (1996), The Hidden Connections (2003), and The Systems View of Life, with Pier Luigi Luis (Capra & Luis, 2014). As well as telling a history of this still emerging discipline, Capra introduces three theories that might serve as its foundation. These are: Ilya Prigogine’s theory of dissipative systems (e.g., Prigogine & Stengers, 1984; Prigogine, 1997), nonlinear mathematics or dynamical systems theory (DyST) (see Abraham, 2011), and Maturana and Varela’s theory of life, cognition, and language, most widely known as autopoietic theory, but by its authors as the Biology of Cognition (BoC). The first two of these remain foundational to what might be called mainstream complexity science. However, Maturana and Varela’s oeuvre failed to be fully drawn into the mainstream of any science, natural or social, and remains as little more than a largely unexplored curiosity, except by a few adventurers. The principal objective of this article is to attempt to rehabilitate the BoC in the context of an evolving epistemology of complex systems, especially the emerging field of the philosophy of complexity.
The paper is presented in two parts. The first providing a brief overview of the history of the BoC, considering its scope and also the features that overlap and resonate with the emerging philosophy of complexity led by Paul Cilliers (e.g. 1998) and his supporters (e.g., Woermann, 2016; Preiser and Woermannn, 2016 and 2018), and based on the writing of philosopher Edgar Morin (e.g., 2008). However, as might be expected there are also significant and profound differences between the two. The one that challenges the current thinking about complex systems most strongly is the claim that organisms are operationally closed self-organizing systems, which are also open to matter and energy. Even more challenging is that Maturana and Varela do not countenance any role for information in the formation of life or the operations of living systems. This position is antithetical, not only to current theories of complexity, but also to the wider established scientific paradigm, from physics and biology through to language theory, and so a robust justification is required. To deal with this issue requires a deeper dive into the BoC, which is done in the second part of the paper, in which a model of an organism-medium system will be introduced, developed, and used to show the mechanism of living without reference to information processing, and which will also be used to demonstrate how the characteristics of complexity are generated through its operation, making it a fundamental building block of complex systems involving life, including human social systems.
Part One
A Challenging beginning
Maturana and Varela’s position on the periphery of the scientific mainstream is well recognized, and various reasons have been given for it. One is the opaqueness of Maturana’s style of writing (Imoto, 2011; Bunnel, 2008, Imoto, 2011). On the other hand, Maturana and Varela’s joint effort to popularize their theory—The Tree of Knowledge of 1992, is easily accessible, as is Varela’s own output (e.g., Varela, 1979), and that with Evan Thompson and Eleanor Roche (Varela et al., 1991). Another reason, perhaps more likely, is that the BoC is a challenge to scientific orthodoxy, the status quo, the established paradigm, being a rare example, if not the only, of the explicit application of second order cybernetics, an approach to knowledge and understanding that recognizes, and accounts for, what the currently dominant paradigm ignores, which is that all explanations are made by, and from the point of view of, an observer to another observer (Maturana, 1978).
Being a peripheral theory, as Heylighen and Joslyn (2002) point out, supporters are too few, and poorly organized to push the theory forward to the main stage. These authors also offer an opinion, that: “It must further be noted that the sometimes ideological fervor driving the second-order (cybernetics) movement may have led a bridge too far. The emphasis on the irreducible complexity of the various system-observer interactions and on the subjectivity of modelling has led many to abandon formal approaches and mathematical modelling altogether, limiting themselves to philosophical or literary discourses.” This is very probably a reference to Maturana and Varela and is a demonstration of how their work is easily dismissed. Their emphasis on the role of the observer in the making of scientific explanations makes their approach open to the wholly unjustified accusation of solipsism, the view the self is all that can be known to exist. Heylighen and Joslyn go further to suggest; “[S]ome people feel that the second-order fascination with self-reference and observers observing observers observing themselves has fostered a potentially dangerous detachment from concrete phenomena.” However, relatively recent developments by Paul Cilliers and others in the philosophy of complexity, to which project Heylighen has contributed (Heylighen et al., 2006), are based firmly on the acceptance of both the observer and the irreducibility of complexity and so provides a hope for the BoC to be reconsidered and possibly rejuvenated as a contributory theory of complexity science.
The Biology of Life, Cognition, Language—and also complexity
Complexity is not something Maturana and Varela address explicitly, but the objects of their study, living systems, are complex system archetypes. The BoC is a theory of complexity. It not just about autopoiesis, but also embraces cognition, language, humanness, emotions, even artificial life. The grand claim of this paper is, as Capra recognised, that Maturana and Varela’s ideas could be an invaluable addition to the current debate on complexity, and make a difference to its direction of travel. Here an attempt is made to present the BoC as a transdisciplinary science, and to place it in the context of some emerging approaches to understanding complex systems and in doing so reveal some complementarity, but also some fundamental differences. The focus will, at all times, be organisms, as exemplary objects of complexity science.
Transdisciplinarity
The emerging paradigm of complexity will be transdisciplinary (e.g., Montuouri, 2013; Jorg, 2011; Bastardas-Boada. 2016). Preiser and Cilliers (2010) suggest transdisciplinarity is embedded in the complexity theory of Morin (2008), and therefore in theirs. The term referring to “a methodology which gives us a conceptual tool to think together those ?elds of study that seem to be situated in opposing positions within the broader scienti?c context.” The assertion in these pages is that the BoC is such a tool, or maybe the toolbox. The BoC is a product of Maturana’s involvement in studies of visual perception in animals (e.g., Lettwin et al., 1959; Maturana and Frenk, 1963, 1964). From the neurological basis of perception, it grew quickly into a comprehensive theoretical framework providing explanations of life and cognition (Maturana, 1971; Maturana & Varela, 1980), language as a biological phenomenon (e.g., Maturana, 1978), biological autonomy, information theory and the mathematics of autopoiesis (Varela, 1979), intelligence (Maturana & Guiloff, 1980), knowing and understanding (Maturana & Varela, 1992), sociality, humanness, ethics and emotions (Maturana & Verden-Zoller, 2008), the nature of explanations and scientific enquiry (Maturana, 1988, 1990), the anatomy and operation of the brain and nervous system (e.g., Maturana et al., 1995), artificial life (Varela, 1997), evolution by structural drift (Maturana & Mpodozis, 2000). Francisco Varela went on to develop concepts such as the embodied mind, and the enactive theory of cognition with philosopher Evan Thompson and psychologist Eleanor Rosch (Varela et al., 1991), and founded a new scientific discipline of neurophenomenology (e.g., Varela, 1996a, 1996b) to explore the nature of mind and consciousness. Varela died in 2000 and Maturana in 2021 leaving a legacy of transdisciplinary research which clearly demonstrates its wide applicability and relevance beyond the headline concept, namely autopoiesis, which has often been misunderstood when applied to social systems (Maturana, 2014).
As previously mentioned, not least of the challenges Maturana and Varela posed to the scientific community at large is an approach to knowledge and understanding that recognizes that all explanations are made by an observer to another observer. To quote Glanvile (2008): Second order cybernetics is: “when Cybernetics is subjected to the critique and the understandings of Cybernetics. It is the Cybernetics in which the role of the observer is appreciated and acknowledged rather than disguised, as had become traditional in western science: and is thus the Cybernetics that considers observing, rather than observed systems.”
To fully explain how scientific explanations are constructed requires a biological explanation of the observer. Hence, to a large degree, the BoC explains what it is to be human: a requirement that current approaches to complexity theory have yet to fully grasp and deal with. Doing so not only strengthens second order science’s claim to be a science of complexity, but also leads to a greater understanding of its relevance to social, organizational and managerial science, and the humanities (e.g., Bond, 2003, 2004, 2005, 2009).
Complementarity with Emerging philosophy of Critical complexity
Complexity science was once feted as the new theory of everything. If this were to be so then social scientists would have to play their part in its development. Paul Cilliers, engineer turned philosopher, emerged to fulfil the role, and did so admirably until he suddenly passed away in 2011. Building upon the writing and ideas of a new complexity paradigm anticipated by Edgar Morin (e.g., 2008), in less than a decade he had established a center for complexity at Stellenbosch University from which he and his co-researchers, principally Rika Preiser and Minka Woermann, began to explore the links between complexity and postmodern and poststructuralist philosophy. This new approach to investigating complex system will be referred to as critical complexity (Preiser et al., 2016). Central to the critical complexity project, is the observer, sometimes explicitly and others implicitly, which provides a point of correspondence from which Maturana’s and the ideas of Cilliers et al. can be compared. The two theories resonate strongly in parts, and where they don’t it is felt due to the underdevelopment of the observer. Here are some areas of commonality.
The observer and Critique of scientfic method
The radicalism in Maturana and Varela’s work stems from a deep and extensive enquiry into the nature of (complex) living systems and acceptance of the observer and of observing, which leads to the following suppositions.
First, we observers make sense of the world—literally, through our sense organs. We do not have direct access to reality, and therefore cannot tell the truth of it and certainly not objectively. This is not to deny reality, but merely to recognize that it impinges on the observer’s (our) senses to induce experiences. Understanding this position, Maturana, as a biologist, as a scientist, develops a way of doing science that is more honest. In this he implicitly accepts that science goes through shifts in paradigm, as theories change practices change, but only when there is consensus. Science is based on agreement, and progresses through ‘compelling arguments’ (Maturana, 1988). Science has never, actually, progressed on the basis of objective truths about reality, but on the agreement that this was the goal of scientists, and there was an agreed and well-established method to achieve it. Because it was so deeply ingrained in practice, it was simply referred to as the scientific method. It nowadays has the label reductionism. The reductionist method enables understanding by breaking apart, by decomposing the object or phenomenon of scientific interest into the smallest possible elements in the expectation that understanding each element separately, their properties, their individual functions, would allow the whole to be understood. For reductionism, the whole was the sum of its parts. As has now become clear, some kinds of system are not open to this kind of approach. There were some systems that are more than the sum of their parts, where properties or qualities, such as life, emerge only at the whole system level. These were designated complex systems and became the object of complexity science, which is widely assumed to embrace an antireductionist or holistic philosophy. As has been inferred above, reductionism also accepts the belief that the scientist, the knower, the subject, the observer, has direct access to concrete reality. Edgar Morin’s position, which is taken up by Cilliers et al., is that complexity science, as it is widely practiced, makes the same presumption and that whatever knowledge is generated through the study of complex systems will be the objective truth. The mind-set remains reductionist, and although the behavior of complex systems is accepted, by definition, as difficult to understand and explain, given sufficient time and resources, they will be explained. This, according to Morin, is to treat truly (really) complex systems as just complicated ones, and is to simplify and therefore to reduce. This approach to complex systems is reflected in a statement by Mitchell (2009:301), who, anticipating the emergence of a new complexity based paradigm, says what is required is the right conceptual vocabulary and the right mathematics with which to “…understand, predict, and in some cases, direct or control self-organizing systems with emergent properties. “ Complex systems, such as living entities, of course are self-organizing. The goals expressed by Mitchell exemplify what Morin calls the restrictive approach to complexity, in particular the aim of controlling. To truly control a system requires that everything is known about it (Bohn, 1994), which is not possible for ‘really’ complex systems. Additionally, Morin says reductionist science eliminates the enquirer, or removes the knower from the process of knowing. It is the goal of second order cybernetics to bring them back. Morin also points out there are truly complex systems that are beyond explanation, that is, beyond the observer’s capability to explain by any scientific method, including the so-called antireductionist or holistic methods, and, furthermore, require a whole new scientific approach to understanding, which he labels general complexity. The principle at the heart of general complexity is that typical scientific explanations cannot be made for complex systems, as the interactions between components are, essentially, impenetrable. Further detail of the notions of restrictive and general complexity can be found in Woermann et al. (2018). The general complexity method accepts that truly complex systems are irreducible to their parts, and, crucially, that descriptions and explanations are made by subjective observers (also referred to as knowers or subjects). As Woermann (2016:100) says: “[O]ur inability to engage with complexity as such is due to the fact we have no direct access to reality. The subject cannot be separated from the object, and nor can the observer be separated from her observation.” With subjectivity arises a supposed problem for scientists, namely—choice. The new complexity paradigm anticipated by Morin will have to cope with both complexity and subjectivity, and therefore choice, and what influences the decisions made by observers/knowing subjects. Thus, issues arise around individual responsibility and ethics, a significant theme in critical complexity, and a topic also explored by Maturana (Maturana & Verden-Zolller, 2008) Subjectivity, therefore, complicates, even complexifies, science.
Alternative explanatory paths
Although Maturana and Varela had no stated interest in complex systems, they were, implicitly, the objects of their studies. The BoC is, essentially a study of complexity, which, in particular, resonates with Morin’s concern for ensuring an appropriate method of investigation. Maturana (1988) in a paper entitled ‘Reality: The search for objectivity or the quest for a compelling argument,’ Maturana pens a critique of the conventional scientific method contrasting two approaches, one that claims it has direct access to reality, the other that doesn’t. These are referred to as explanatory paths, which are very awkwardly entitled, objectivity-with-parenthesis (‘objectivity’), and objectivity-without-parenthesis (objectivity). Objectivity (without parenthesis) equates to the reductionist method. Objectivity (with parenthesis) simply recognizes that observing is subjective. Accepting this does not lead to a devaluation or diminution of science, it merely clarifies how scientific knowledge has been, and can only have been, generated, and highlights the evidence that science progresses through ‘compelling arguments’ and through revolutions in thinking, as Thomas Kuhn has shown. How to cope with this realization is, in this author’s opinion, the essence of Morin’s complexity paradigm.
Relations and parts
It is claimed that by focusing so strongly on decomposition and the separation of parts, the relationships that make the whole more than the sum of its parts are lost. “In ‘cutting up’ a system, the analytical method destroys what it seeks to understand.” (Cilliers 1998:2. See also Woermann et al., 2018). Although Maturana and Varela see no way around ‘cutting up’ a system by drawing distinctions, as this is how we sense the world, they do most strongly adhere to the importance of relationships, which is evidenced in the centrality of structure and structural change in the BoC. For example.
‘A system is a collection of elements that interact and relate with each other in such a way that the interactions that any of those elements have, and the results of these interactions, depend upon its relations with the others.’ (Maturana, Mpodozis & Letelier, 1995)
Organism and environment
The relationship between organism and environment is a common point of focus in many disciplines, and is most prevalent in theories of biological evolution, Darwin’s evolution by natural selection being the most prominent. There are many others, including Maturana’s ‘evolution by structural drift’ (Maturana et al., 2000), in epigenetics (Jablonka & Lamb, 2005), also known as the extended evolutionary synthesis (Pigliucci & Müller, 2010). Theories subsumed under these new labels include: niche construction theory (NCT) (Laland et al., 2000), and Developmental Systems Theory (DST) (e.g., Oyama, 2000; Oyama et al., 2001). Also concerned with the relations between organism and environment are so-called, extended theories of ‘the mind,’ which focus on the role of the environment in, as do more classical theories of psychology and perception: James Gibson on visual perception (2015/1986), Gregory Bateson’s work on environment, cognition, and the mind (1980, 1972/1987). However, all these ideas have been developed on a path parallel to the BoC and second order science, and are not underpinned by the same reasoning, they follow the explanatory path of objectivity (without parenthesis). This is also the case for critical complexity when it comes to explaining the relation between organism and environment.
Morin-Cilliers et al. recognize the co-dependency of complex system and environment. Both BoC and critical complexity recognize the openness of complex systems, the significance of boundaries, and that openness is the key to understanding how they exist and persist together, as all evolutionary theories require. Both also agree that both energy and material is exchanged. However, in critical complexity, the explanations of co-existence, or co-dependency, remain consistent with a reductionist paradigm, especially its adherence to the information theory of Shannon (1948), Shannon and Weaver (1963). Cilliers, for example, says: “In order to respond appropriately to its environment, a complex system [such as a living entity] must be able to gather information about that environment and store it for future use.” See also Cilliers (2016) on the operation of the information processing brain (pp 23-38). Similarly, “…open systems can only be understood in terms of their relation with the environment. This is because there is an energy, material, or information transfer into or out of a given system’s boundary.” (Woermann, wat is comp paper ). The inclusion of information is totally rejected by Maturana and Varela and is the point where the two approaches begin to diverge.
The primary reasoning for the rejection of information arises from the idea of operational closure, which is firmly connected with the whole notion of living systems as self, or spontaneously organizing, self-making (autopoietic) systems. To fully justify the rejection of information and the acceptance of a closed but open systems will require a much more in-depth understanding of the principal concepts underpinning the BoC, to which the remainder of the paper is dedicated, barring final comments. Cross referencing to critical complexity will be made as appropriate.
The operatonal closure of an open system
What has proven to be one of the biggest challenges is the apparent paradox that an organism is at once both a closed and open system. This section will address the issue, which will also be a vehicle for introducing some of the key words and concepts used by Maturana and Varela.
The operationally closed but open living system
Some time in the deep and distant past life began when:
[A] living system arose in the history of the earth in the moment in which some network of molecular productions became closed upon itself constituting a discrete entity in which the molecules produced realized the same network of molecular productions that produced them while realizing at the same time its boundaries as an autonomous totality in a molecular medium with which it was in molecular interchange. (Maturana, 200:7)
Maturana is here referring to the operationally closed process he named autopoiesis. The challenge to current scientific orthodoxy is contained in three words ‘closed upon itself’ which process, under the right conditions, can be referred to as spontaneous organization (essentially self-organizing), but also described as natural boot strapping, or the loop of life (see Maturana and Varela, Chapter 2, and Brockman, 1995, chapter 12). However, the loop is not an endlessly repetitive molecular production process. Instead this circular operation is, very significantly, recursive, and irreversible, and involves a network of processes producing molecular components that spontaneously interact to regenerate the network of processes that produced them. This is a cycle in which nominal components are continuously being transformed, including the cell wall, which is not simply a container for processes. Single cells are autopoietic, and of the first order of autopoiesis. Multicellular organisms are second order, and collections of multicellular organisms that operate symbiotically at the molecular level are third order autopoietic systems. Social systems are not autopoietic, as Maturana argued against Niklas Luhmanns insistence to the contrary. The strength of the challenge is further magnified as the brain and nervous system, and immune system, in higher order life, is also operationally closed. The fullest implications are only explored briefly below.
The question is, of course, if living systems, i.e., organisms, are closed upon themselves, how is the very necessary need for molecular interchange between organism and ‘environment’ achieved, which it evidently is? While it is important to understand organisms are operationally closed systems, it is equally important not to confuse this idea with the thermodynamic notion of closure, meaning a system is totally isolated from its surroundings with no exchange or transference of matter or energy. While both Varela (e.g., Varela, 1978 and 1979) and Maturana (2002) have been careful to concede that claiming a system to be open and closed at the same time is somewhat paradoxical, they have also offered an alternative mechanism based on their notion of structure, which is presented with a model organism-medium system presently. Woermann (2016) is aware of the closed-open paradox, , although relies on a secondary source (Hayles, 1994, 1999), who “criticizes the idea of organizationally-closed systems, and remains unconvinced that Maturana and Varela are able to solve the problem of solipsism.” (Woermann, 2016:33). Woermann appears to accept this. However, she revisits the issue of closure and openness from time to time. She says (Chap.7.) Morin has criticized the rigid opposition between open and closed systems, arguing that complex systems are neither completely open or closed, but neither discover an answer to this paradox.
With regard to exchanges of matter between organism and medium, this is easily dealt with. Operational closure is predicated on the basis that circular or cyclical metabolic processes are continuous, on-going. Individual molecules are assimilated, taken in by, or absorbed, but they are not regarded as inputs. In general, inputs are considered to trigger changes in the system, often they are conceived as instructions to start. However, what happens within an organism is determined by its structure, moment by moment (further explanation below). If organisms were considered to have to receive a constant stream of instructions to keep going, they would not be self-organizing. The acceptance of operational closure goes hand-in-hand with rejecting instructional inputs, which are those that trigger, or programme for, behavior of any kind, including energy and information. It is implicit in most input-output modelling that inputs are of this kind, but then there’s information.
(T)he nervous system does not 'pick up information' from the environment, as we often hear... The popular metaphor of calling the brain an 'information-processing device' is not only ambiguous but patently wrong. (Maturana & Varela, 1992:169).
Why is it wrong, when most scientists believe it to be true? Two main reasons are offered. First, the more obvious one, information is immaterial; it’s a concept and cannot be processed by a concrete physical system. Belief that it can begins with an all-too-common error, found in many scientific explanations, of conflating what Maturana and Varela have called phenomenological domains. These will be dealt with before considering information in more depth, and use will be made of a model of the organism medium system, but first the definition of medium.
The medium
As with many other familiar terms, environment, medium and also niche take on slightly different meanings and function in the BoC as compared to common usage. From an observer’s point of description, the environment is the context in which an organism is distinguished. Both medium and niche, on the other hand, are specified, as far as an observer can do this, from the point of view of an organism.
A medium is that part of a conceived environment in which an organism operates and which, therefore, supports its existence. It presents opportunities for interactions that maintain a particular manner of being in the world, but also events that may jeopardize a life and therefore induce avoidance behavior. A medium is unique to a specific organism. E.g., spiders operate/live in a different medium to an ant, or an ant eater, but from an observer’s normal point of view they all live in the same environment. Note also that organisms have senses that are ‘tuned’ to their respective medium; indeed they may not be capable of distinguishing the medium of another creature. For example simple forms of life exist in a molecular medium. For many, if not most readers, this interpretation of medium will equate to the idea of a niche, which, in the BoC, also has a different meaning.
Niche is a space (or domain), contiguous with the nominal boundary of an organism, where, it might be said, physiological processes extend into the medium (after Scott Turner, 2000, 2004). Life sustaining physico-chemical change occurs within the niche they specify. With respect to we humans, for example, these include oxygen/carbon-dioxide exchange within the lungs, the breakdown and transference of life sustaining molecules aided by enzymes and bacteria in the gut, and also the reactions at the sense organs with light, heat, sound, and smell (molecular interaction). The niche also places physical constraints on behavior. For example, we humans cannot go where the atmosphere is ‘thin,’ although some of us are well adapted to the high life. Very importantly, for living entities the niche marks the boundary, albeit a fuzzy one, between what Maturana and Varela refer to as phenomenological domains.
The organism-medium system (OMS) and phenomenal domains
The organism-medium system (OMS is roughly equivalent to what Morin-Cilliers et al. refer to as complex system-environment, but further detail will follow shortly. There are two phenomenological domains, the domain of behavioral phenomena, and the domain of physiological phenomenon. An observer’s primary point of view is always the behavioral domain, which most would construe as the observer’s environment. Descriptions of the interactions that are observed to take place in this domain refer to system level properties and behaviors. System level properties include the wetness of and fluidity of water, the roughness of sandpaper, the shape of a hammer, the hardness of steel. Qualities of animals and humans, such as emotional state, can be judged from their behaviors. The tiger acts in anger, the man behaves intelligently. In such descriptions the individuals, the unities, are treated as wholes. In other words, the organism part of the OMS is treated as a whole and interactions described in terms of whole system properties. Organisms with nervous systems are observed to act on their environments, but in describing that behavior and its result(s) an observer would not normally attempt to decompose the organism. So for example, consider making a description of hammering, as when a blacksmith hammers a bar of hot iron into shape. There is probably no urge to decompose the blacksmith or refer to the microstructure of the metals of hammer and object of its action, but there might be a tendency to infer the goals or the motivations of the blacksmith.
The term hammering is also used to describe what happens to the heart, when someone receives a fright, or falls in love. However, they are not acceptable scientific descriptions of what happens in the domain of physiological phenomena. At best these types of descriptions are metaphorical. With regard to constructing scientific explanations, it is a fundamental error to express what happens in the domain of physiological processes in the same terms used to describe interactions in the domain of behaviors.
The appropriate form of explanation of physiological phenomena, such as the operation of a living system, is referred to as a mechanical or mechanistic explanation. This is not equivalent to treating an organism as an allopoietic machine, one to which the input-output form of modelling is best suited. Maturana explains as follows, which is an echo of the Morin-Cilliers et al. position vis a vis their preference for emphasizing relationships rather than components .
In a mechanistic explanation, the observer explicitly or implicitly accepts that the properties of the system to be explained are generated by relations of the components of the system and are not to be found among the properties of those components. The same applies to the mechanistic explanation of a phenomenon, in which case the observer explicitly or implicitly accepts that the characteristics of the phenomenon to be explained result from the relations of its constitutive processes, and are not to be found among the characteristics of these processes. (Maturana, 1978)
Note also that the nature of the medium is very different in the behavioral and physiological domains. For example, in a living system, the individual cells, exist in a medium consisting of other cells and the molecules exchanged through the process of autopoiesis, recalling that every cell is autopoietic and, as indicated later, the cell-medium system is complex in its own right.
The error of conflating domains is common in many a scientific treatises. For example, if one was to describe the interactions of the immune system in terms of attack and defense, or the operations of the brain as if it were a computer. These are also examples of what Gantt et al. (2012) termed metaphors turned explanations. In other words, the metaphors are so ingrained that their origin in descriptions of behavioral phenomenon simply gets lost. It is a feature of language that metaphors often get so subsumed in everyday speech that they just disappear from awareness. Language, it appears, is built on a reef of dead metaphors (Deutscher, 2005).
Brain-nervous system as an operationally closed and complex system.
Operational closure of organisms is controversial but rarely does a commentator on the topic venture into what is potentially an even more controversial claim by Maturana and Varela’s that the brain-nervous system, which shares the sense organs with that which embodies it, is also operationally closed (but also open to matter and energy). There are significant and profound implication that follow this claim, but space does not allow further exploration. One consequence is that neurons perturb other neurons, via a molecular medium. It should also be appreciated that the neuron-medium interaction is also similar to that proposed for the organism-medium, it is a recursive process and therefore generates complexity (see below).
Information
In his foreword to Morin (2008), Mutuouri recognizes that, in complexity science, information establishes a link with physics, “…while at the same lime being fundamentally unknown in physics.” “Information is a problem concept, not a solution concept. It is an indispensable concept, but it is not yet an elucidated or elucidating concept.“ (Mutuouri, 2008:14). To the contrary, Varela (1979) claims it is dispensable. Likewise, Mitchell sees problems with the concept of information and its processing by living systems, saying: “In many people’s minds information has taken on an ontological status equal to that of mass and energy—namely, as a third primitive component of reality. […] However, like several other central terms of complex systems science, the concept of information processing tends to be ill-defined; it’s often hard to glean what is meant by information processing or computation.” (Mitchell, 2009, p169).
There is an oft-referenced definition of information: that which informs. To be informed, is to be shaped, or changed (Bob Doyle the self styled information philosopher. Found at informationphilospher.com). Similarly, Varela uses the term in-formare, meaning to form within. In an attempt to clarify his position, Varela (1979:266) makes a decision to distinguish between information and in-formation. The former carries the conventional meaning and refers to something or someone; or it’s an instruction, a representation or symbol, a mathematical formula, for example. In-formation is something constructed within the observer, who feels shaped, changed by it. This reference to shaping, forming, or the changing of a receiver is perfectly in keeping with Shannon and Weaver’s famous model of communication (or information theory). If the same message is sent repeatedly to a receiver, he or she ceases to be in-formed after first receipt. After that there’s nothing to distinguish, nothing new to perturb the receiver, nothing else to change or shape them, or respond to. The only way an observer can tell when a receiver of a message has been shaped or changed is by their behavior, which is conceived as part of the process of information processing. Being informed arises in the behavioral domain. The inference that someone processes information, is just that, a guess, and is not a physiological explanation. An observer-receiver, then, is in-formed, but never receives an input of information that determines their behavioral response.
This necessarily brief discussion of information barely even scratches the surface of the issue, but hopefully the reader can conceive of the possibility that information as a ‘third component of reality’ is dispensable. The idea that living systems receive and process information is due to an error of conflating domains, or describing physiological processes in terms appropriate only to the behavioral domain. Having concluded thus, the question that now needs to be answered is by what mechanism does the observer feel in-formed, changed within? The answer is in ‘structure’ which matter will be addressed after considering the organism-medium system as a unit of complexity.
The Organism-Medium system as a unit of complexity
According Jim Meiss (2003),1 a complex system is a spatially and/or temporally extended nonlinear system characterized by collective properties or behaviors associated with the system as a whole, which are different from those of the constituent parts. Individual organisms appear to match this definition. Life appears to be an emergent property of a whole organism. However, more careful observation reveals that behavior is not a property of an organism alone, but of the organism-medium system.
Behavior consists of various different actions or events that are distinguished by an observer. Particular instances of behavior are explained as a particular response to a particular cause, which makes them effects or results. Living, as a process, is marked by behaviors, which are responses to changes in the medium in which they exist with other similar and different entities. Maturana and Varela, also Morin, recognized that life and living could only be explained if the environment was incorporated into the explanation. The biologist’s solution, mostly implicitly, was to treat the two—the organism and the medium—as one system, which then becomes the fundamental unit of analysis of a theory of living. It is suggested that the OMS is also the source of complexity in all systems involving living entities.
Part two
A model of the complex organism-medium system (OMS)
What is proposed here is a model of the organism-medium system that will be used to illustrate how the two persist together without reference to the processing of information. At the same time, it will also be used to show its operation as the source of complexity in all kinds of social systems. A modified diagram suggests cell-medium interactions will be complex too. For the sake of clarity and ease of explanation the primary assumption is that the model refers to an organism with a nervous system able to move through and experience its medium, which includes all observers.
The basics: System dynamics
The initial approach to modelling is simple, and is nominally based on the widely used systems thinking method developed and applied by the management and system theorists Daniel Kim (1990) and Peter Senge (2006). The rules for systems thinking are drawn from systems dynamics (SD) (e.g., Wolstenholme, 1990). Referring now to the model in figure 1, the immediate purpose of which is to show that changes in organism and medium are reciprocal.
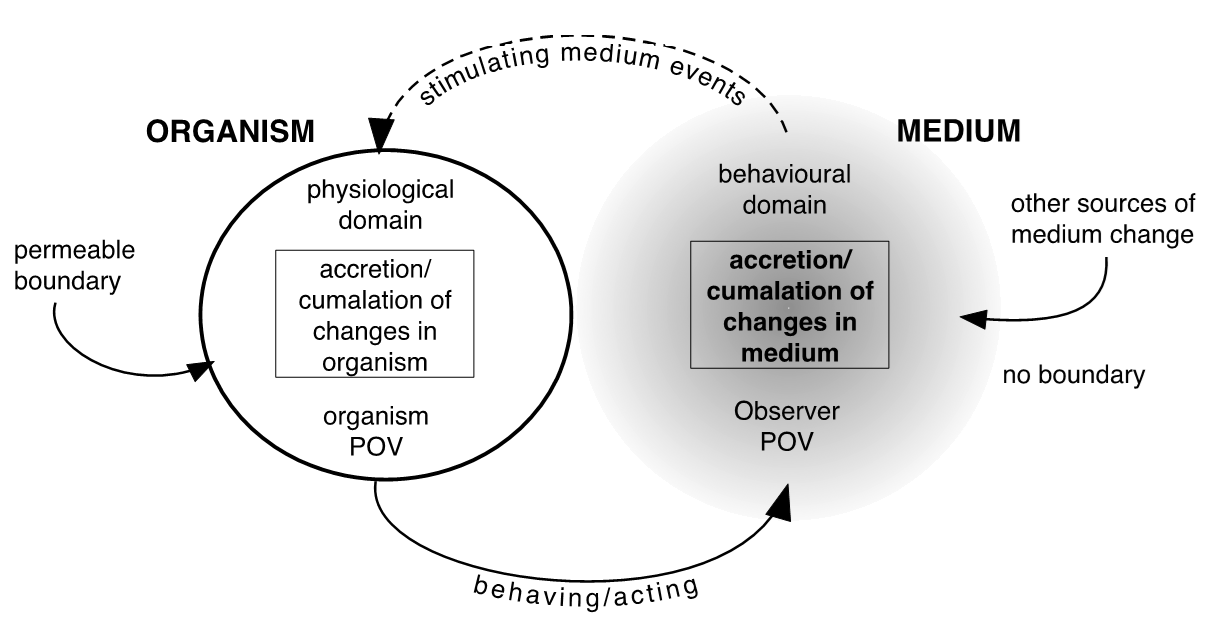
Figure 1. The generic model of the organism-medium system.
The kind of scenario that this model represents is any kind of routine living. It can represent a pet dog or its owner walking through a woodland glade. First, the organism is delineated with a circular line indicating a permeable boundary plus its niche. The boundary is a component of the organism, and is permeable, but not to instructive inputs. The medium is treated as unbounded. Although the choice of modelling approach restricts this being shown, the organism, of course, exists within its medium. The diagram shows a circular relationship between the two. Although suggesting sequential changes, the cycle will take nanoseconds. Changes occur moment-by-moment. Changes to the medium are results of a process labelled ‘behaving’ or ‘acting,’ suggesting the organism physically acts on the medium to change. The simple act of walking crushes whatever is underfoot, and dogs defecate. Changes to the organism are said to occur through perturbing. The latter requires some further knowledge of Maturana and Varela’s concept of structure before it can be explained satisfactorily, but for now consider changes in the organism are induced or stimulated by events experienced within the medium. Staying with the woodland walk. These can include smells, a shift from shade to sunshine, a scratch from brushing by a thorny bush, the bark of the dog, the dog hearing the sound of whistle blown by the owner. In SD modelling, change is typically expressed in numbers of units, be it cars, population, or bottles of beer, but changes in qualities of a starting item (e.g., organism or medium) can also be accommodated and are here indicated by use of the terms accretion and cumulation. Accretion refers to a gradual change through a layering process i.e., one change on top of another, while cumulation refers to a gathering and combining of results, so producing an overall change in qualities or attributes. Maturana refers to this process of layered changes as epigenesis. Also indicated in the model are the two phenomenological domains—behavioral and physiological, as two points of view, the observer point of view (POV) and an imaginary point of view within the organism that will be useful to consider later. The processes and results of will be considered in the next section.
Perturbing and inducing change
Even with respect to everyday events and behaviors, we observers are constrained in the form of explanations we can make. Each of us tends to conceive a particular kind of behavior as a response to a particular cause as an input that triggers it. We are so used to falling back on inputs and outputs of information to provide an explanation of how systems operate that it is difficult to conceive of an alternative, but the idea of perturbing is one.
In physics a perturbation is defined as a slight alteration of a physical system caused or induced by a secondary or indirect influence. This is the meaning used in the BoC to describe what happens within an organism in response to non-destructive external events it encounters through its senses. From our human point of view, to be perturbed is to become conscious or aware of changes, to distinguish them, to feel them. Biological sensory instruments are integral to both the brain-nervous system and the body that envelops it. Changes in so-called receptor neurons are induced by energetic stimulation by light, heat or electricity, or by molecular interactions in the senses of smell and taste. The general mechanism is one of induced restructuring, or reconfiguration of an organism’s structure. Perturbation is what happens when someone is in-formed. In higher animals, especially humans, moment-by-moment reconfigurations of the brain-nervous system are by far the most consequential. However, simpler organisms without the latter, still sense interactions with their medium. E.g., when a bacterium is placed in a sugary medium it senses the concentration gradient and moves along it (Maturana & Varela, 1992:149-150, and Thompson, 2011).
When an organism is perturbed, through its senses, structural changes are induced, which can also be felt/distinguished as an emotion. Reading an unexpected card from a distant admirer on Valentine’s Day might produce a pleasant change within, as in being in-formed. An emotion is not something abstracted from experience, but is a configuration of structure that Maturana (1989) defines as a disposition to act. To further understanding the process of perturbing it is necessary to delve a little deeper into autopoietic theory, and consider what Maturana and Varela say about structure, and its corollaries, structural coupling and structurally determination.
The BoC Basics: Structure and its corrollaries
Structure
Generally speaking, structure is considered to involve at least semi-permanent and rigid relationships between parts. Within the BoC, however, structure is fluid, as is hopefully demonstrated here. First consider what happens in this domestic accident.
This morning I approached a kitchen cupboard with the intention of taking out my favorite breakfast cereal. I opened the door, reached inside, grasped the box, withdrew my arm, and then, quite unexpectedly, the cuff of my shirt caught on a protrusion inside the cupboard (part of the means of fixing the handle/knob). The sudden snaring of my cuff caused my arm to jerk as I drew it out of the cupboard. The jerking caused me to lose my grip on the box. Momentum propelled the now freefalling box out of the cupboard and toward the floor. On its way, it hit the edge of the kitchen worktop, positioned directly below the cupboard, turning it upside-down. When the box hit the ground its contents were scattered a meter across the floor.
Superficially, the story tells of a sequence or chain of anticipated and unanticipated results. Grasping the box was intentional, but catching a cuff on the internal protrusion was not, nor was losing grip of the box or it hitting the worktop on the way down before the contents were scattered. Note, as each result occurred, a different set of components was brought into play. Unexpected results began when the shirt cuff connected to the protrusion on the cupboard door, which connected it, indirectly, to the box. This was followed by the connecting of the worktop with the box, and then the box with the floor. Although the story explicitly relates a series of results, it is as much about the formation of a series of different transitory structures marked, located in space and time by results distinguished by an observer. At a particular moment, a particular configuration of a particular set of components came into being to produce a particular result. Thus, structure and results are intimately entwined.
In the BoC, structure is defined in different ways, e.g., “the presence and interplay of components in a given space” (Maturana, 1978). Clearly this is pertinent to the above illustration. Or, in relation to a system: “[T]he structure of a particular [system] is the manner in which it is actually made by actual static or dynamic components and relations in a particular space…” (Maturana, 1988b:6.iv.) Alternatively structure is defined as: “The actual relations which hold between the components which integrate a concrete machine [or system] in a given space.” (Maturana & Varela, 1980:138). This leads to the definition of a system used earlier. “A system is a collection of elements that interact and relate with each other in such a way that the interactions that any of those elements have, and the results of these interactions, depend upon its relations with the others.” (Maturana, Mpodozis & Letelier, 1995:16). Note no reference is made to inputs or outputs, only results. There’s no reference, either, to system purpose, which is common to many definitions. From the description above, the operation of a system can be conceived as a continuous transformation of relationships between objects, which are only system components for fleeting moments, and that, in those particular moments, the actual relations between the actual components of the system produced a particular result. It could be said that the nature of the components, e.g., their shape, material constitution, and surface quality, plus the specific relations between them at a particular moment enabled the result. Instead of enabled, one might say that all results were determined by the nature and relationships of the individual components, but at a particular moment in time. It might also be said the outcome arose spontaneously from the structure formed the moment before the result. In which case, no other outcome was possible from that particular structural configuration. This is what structural determinism is and it applies to every kind of system, natural or designed and engineered.
This interpretation of structure is different from the more familiar engineer’s notion of structure in which the relationships between durable components form something rigid, unchanging and enduring and yet it is still forms part of an explanation for the persistence of specific kinds of organisms for planetary significant periods of time, whether they are bacteria, dinosaurs or fungi. Moreover, so long as a particular structure persists, a particular function does also. As illustrated in the cereal spillage, the structure of a system, moment by moment, determines a particular result in that moment. That which is distinguished as the final result becomes associated with the function or purpose of a system. Although structure determines or enables a result, it constrains simultaneously, a principle also recognized by sociologist Anthony Giddens (1984) and embodied in his principle called the Duality of Structure. See also Mingers’s (1996) comparison of the two conceptualizations. That stated, in a further exposition of the nature of structure, Maturana & Varela distinguish between it and organization, which is dealt with in the section of structural determined behavior.
Structural coupling of organism and medium
For organism and medium to persist as one system, correspondence between them must be maintained. The mechanism for this is structural coupling. The medium, which is an independent system (see definition below in section on structural determination), is changed by the actions of the organism. As life evolved, the medium became populated with many other organisms and their actions too became a source of perturbing events. Thus, the medium continuously holds opportunities for new interactions between different kinds of organism. When such interactions are repeated time and again, a structural coupling of multiple organisms occurs and the ontologies of every organism become entwined with every other, whether plant, animal or fungi. When two organisms interact, the structural changes of state in one become perturbations for the other, and vice versa, in a manner that establishes what Maturana (1978) has described as an interlocked, mutually selecting, mutually triggering domain of structural or state trajectories, a process that becomes central to an alternative theory of evolution known as natural (structural) drift. It should also be appreciated that the medium is equivalent to an organism’s world, being a space defined by the history of interactions that sustain its living.
Structural coupling is formally defined as follows.
When two or more structure determined systems enter in recursive interactions and undergo structural changes without losing their respective class identities [organization], their structures change together congruently, and there is structural coupling. We call structural coupling both the dynamics of coherent structural changes that occur in such a case, and the condition of structural coherence that takes place as a result of that dynamics. (Maturana & Mpodozis, 2000:48)
This introduces a further concept, structural determined system, which is addressed following another brief aside about the difference MATURANA AND VARELA hold between structure and organization, which has a bearing on the kind of interactions an organism can engage with.
Organisms and Structure determined behavior
According to Maturana and Varela, an individual organism can only be specified or defined with reference to both its structure and its organization. Although each member of a species might differ in structure, they each have the same organization. The significance of this is that an organism’s structure can change, so long as the organization that defines it as a particular class of organism is not compromised. In other words, there is a limit to allowed interactions. Moreover, structure, being the actual relations between the actual system components, determines the kind of interactions in which a specific organism can be involved moment-by-moment. The formal definition of a structure determined system is:
[A] system in which all that happens happens as a structural change determined in it at every instant by its structure at that instant, regardless of whether this structural change arises in it in the flow of its own internal dynamics, or contingent on its interactions. (Maturana, 1988:36)
The implication of the above interpretation is that structure determines results. As indicated in the cereal accident, the existence of a particular structure is at first inferred from a result. With respect to organism, a result perturbs. A result is distinguished by senses, which act sets up a series of disturbances within the organism. However, structure alone does not determine whether or not an interaction proceeds, energy is also required and this is where Prigogine’s thermodynamic theory becomes relevant. Although the space does not allow for deep discussion on an organism’s as dissipative structures, the requirement for heat energy is clearly evident. Take this simple but familiar form of molecular interaction as an example.
It might be described in terms of behavior, an attack of acid, an infiltration, a penetration of the weak base of calcium hydroxide, which is ripped apart, aggressively denatured, destroyed. Alternatively, it could be said that under favorable conditions of energy availability and proximity, the acid and the base simply undergo spontaneous structural change. Again, no input is required, just spontaneous, even explosive, adjustments to relationships that result in two different molecules.
H2SO4 + Ca(OH)2 = Ca(SO)4 + 2H2O
Sulphuric Acid + Calcium Hydroxide Calcium Sulfate+ Water
It is suggested that it is this spontaneous change of relationships between molecules that occurs within the space of the niche enabling material exchange. What can and can’t be exchanged is determined by structure, the extant relations between actual components of the organism and potential components.
From the discussion so far it might be surmised that structure, rather than autopoiesis, is the core concept in autopoietic theory as it underpins much of how the organism and medium are co-created, and remain coherent. Structure enables what is happening in any instant, but constrains something different happening. This is why structure is said to determine, and determines too what can enter an organism if its coherence is to be maintained. Structure also determines what happens in non-bionic machines. In the definition of structure given above, the term ‘actual’ is a reference to the precise nature of components and relationships. Components have physical shape and qualities. If a steel cog in an industrial machine was replaced by one of the same shape, but made of cheese, it would not run, but a wooden one might enable operation for a time. If the surface of a metal cog was covered in sand paper, it would have different surface properties and again ‘proper’ operation would be hindered. ^ ^ With these pieces of theory in place it is time to examine how the characteristics of a complex system are generated by the O-MS.
Generation of Complexity by the organism-medium system
Recursion
The term recursion has a variety of meanings according to the disciplines in which it is used. The most common applications of recursion are in mathematics and computer science, but also in language theory. In the BoC, recursion is brought into play in the concept of the recursive process:
When a repeating circular process becomes (structurally) coupled with a linear one that displaces the circumstances of the repetition, the repetition of the circular process becomes a recursion, and a new phenomenal dimension appears. (Maturana et al., 1995:17).
Also:
In biological systems, recursion is a fundamental dynamics, because of the circular character of biological processes and the linear character of the relations between a living system and its changing medium. (op cit).
Maturana offers an analogy to help readers grasp the concept of biological recursion, that of a motorcar moving along a road. A similar idea is offered here, a locomotive on a railway, which analogy aids comprehension because it also serves to reinforce the concept of structural coupling.
Recursion occurs when a repeating circular process is structurally coupled with a linear one, which then displaces, in space and time, the circumstances of the cyclical repetition. Movement is the phenomenon an observer distinguishes. Figure 2 below is an attempt to help explain this particular formulation of recursion. Note that the process is irreversible and the ‘circular’ part of the diagram represents a series of separate cyclical metabolic processes that take place moment upon moment, and not a spiral, which would be linear (a metal spring can be pulled into a straight line).
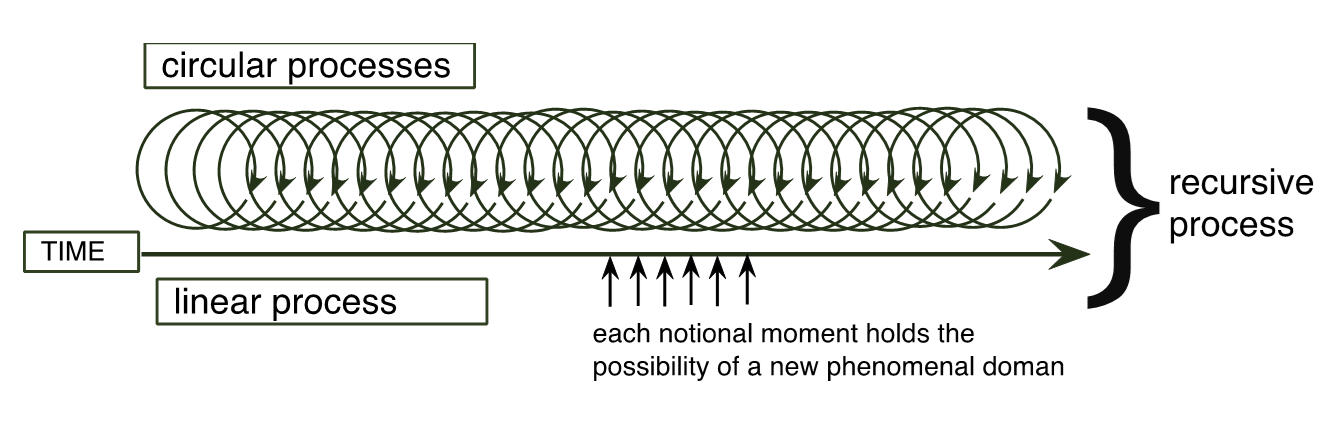
Figure 2. A recursive process.
If the locomotive represents an operationally closed organism, the railway (or track-way) represents its medium of operation with which it is coupled. The loco and the track-way are two independent systems. The railway includes the steel rails, the sleepers and the, mostly unseen, foundation, which in turn is connected to an underlying (also unseen) supportive geological structure. Structural coupling is achieved and maintained through a combination of shape and the material from which wheel and rail are made. The two are not fixed in any other way, and there are no discernable inputs or outputs between them. If a loco’s wheels were somehow hidden from view, then all that would be observable is its linear movement, forwards or backwards. The locomotive moves because, as the relationships between the internal components (engine/transmission and other mechanisms) change, so too does the relationship with the track way with which it is coupled. Moment after moment, the internal structure of the locomotive changes as does the relation between it and the track way. An actual railway (the medium) is not a passive structure, it responds to shifting forces upon it, nor is it ever entirely uniform. With each moment of linear motion, the locomotive’s mass is supported by a slightly different structure. What happens in each moment will depend on the structure of the locomotive, the structure of the railway, which has the potential of changing the efficacy of the coupling. The locomotive will continue to move linearly, because, and so long as, structural coupling is maintained.
An analogy or metaphor only takes understanding so far, and can mislead. A locomotive is not autopoietic, but an allopoietic machine that does not produce itself. The structural changes that occur within an operationally closed organism are adjustments to the structure of the moment before. With regard to individual organisms, structural changes occur on top of previous changes in response to perturbations induced by events sensed in the medium in a recursive fashion indicated by the model of figure 1. Multiple organisms, or multiple cells, are each part of the medium of the other so collections of organisms perturb each other in subtly different ways so multiplying the uncertainty of the totality thus making it increasingly difficult to pinpoint a single or even multiple source of a causal change, which idea resonates with that of Cilliers: “…complexity is not located at a specific, identifiable site in a system. Because complexity results from the interaction between the components of a system, complexity is manifested at the level of the system itself” (Cilliers, 1998). That is to say, complexity is manifest in the behavioral domain of phenomena. Complexity, then, is an experience. Evolution is evidence that structural constraints operating within an organism do not totally conserve, because recursion moves the system in its medium to a different ‘present.’ From moment to moment the medium is changed by the behavior of its constituents thereby potentially changing the kinds and dynamics of interactions. Organism and medium evolve together, as explained by Maturana et al. (2000) as evolution by structural drift.
Concluding remarks
This paper began with a claim that Maturana and Varela’s body of work is of value to those scholars who seek to understand complex systems. The first part sought to find and prove connections to the ideas on complexity that originated in the writing of Edgar Morin but were developed by Paul Cilliers and his colleagues Rika Preiser and Minka Woermann, as critical complexity. The purpose of the second part was to use a model of the OMS try to establish a position from which the apparently serious differences could be remedied. The principal idea to be grasped in the narrative about the OMS is that the behaviors that define a complex system arise in its operation as a structurally coupled, recursive structure determined system, capable of generating unexpected changes within both the organism and its medium. If an observer distinguishes such changes, that is, if they become conscious of them, then they are perturbed and changed by them. They become in-formed, but they don’t receive inputs of information. One very significant manifestation of being in-formed, formed within, are emotions, another area that conformist research tends to dismiss. Although not explored here, a subsequent paper (Bond, forthcoming) will show how significant a factor it is in the dynamics of human social systems. The pursuit of an understanding of emotions leads to the softer side of complexity, and issues around decision-making, responsibility and ethics, that are of special interest to Cilliers et al.. Thus, rises the possibility of a Maturanian perspective on postmodern and post structuralist philosophy so building on the connections already made between the BoC and Kant (Weber & Varela, 2002), Merleau-Ponty (Varela et al., 1990), and other phenomenologists such as Husserl and Heidegger (e.g., Reynolds, 2004).
There is plenty left to explore, Maturana’s claim we humans live in language, or in conversations, their ‘take’ on culture, the ethics of ‘technology’ (Maturana, 2005; Bond, 2003, 2004, 2005), and if the BoC were to be accepted as a ‘complexity science’ then it would bring a fresh complexity flavored approach to these and many other areas of scientific interest.
References
Abraham, R.H., (2011). "The genesis of complexity," World Futures, 67:4-5: 380-394.
Bastardas-Boada, A. (2016). "Toward Complexics as a meta-transdiscipline field," Available at: http://diposit.ub.edu/dspace/handle/2445/63387. DOI 10.13140/2.1.3487.5686. Accessed 22-01-22.
Bateson, G. (1980). Mind and Nature, London: Fontana.
Bateson, G. (1987). Steps to an Ecology of Mind. Chicago, Ill. Univ. of Chicago Press. (Orig. 1972).
Bond, P.L. (2003). "The biology of technology: An exploratory essay," Knowledge, Technology, and Policy, 16(3): 125-142.
Bond, P.L. (2004). "Maturana, Technology, and Art: Is a biology of technology possible?", Cybernetics & Human Knowing, 11(2): 49-70(22).
Bond, P.L. (2005). The emergence of complex emotioning innovating and polytechnical systems. An essay on the biology of technology. Complexity and time workshop. Conference on Complexity and Society, Liverpool, Sept 2005.
Bond, P.L. (2009). "Toward a living systems framework for unifying technology and knowledge management, organizational, cultural and economic change," in D. Harorimana (ed.), Cultural Implications of Knowledge Sharing, Management and Transfer: Identifying Competitive Advantage, Hershey PA: IGI Global.
Brockman, J. (1995). The Third Culture: Beyond The Scientific Revolution. New York: Simon & Schuster
Bunnel, P. (2008). "Editor’s foreward," in H.R. Maturana, G. Verden-Zoller and P. Bunnell (eds.), The Origin of Humanness in the Biology of Love, Exeter, UK: Imprint Academic.
Capra, F. (1990). The Turning Point: Science, Society, and the Rising Culture, New York: Simon and Schuster.
Capra, F. (1996). Web of Life, London: Flamingo.
Capra, F. (2003). The Hidden Connections, London: Flamingo.
Capra, F. (2007). "Complexity and life," Systems Research and Behavioral Science Syst. Res. 24: 475- 479
Capra, F., and Luis, P.L. (2014). The Systems View of Life. Cambridge: Cambridge University Press.
Cilliers, P. (1998). Complexity and Postmodernism, London: Routledge
Cilliers,P., and Preiser, R. (2010). Complexity, Difference and Identity, Springer Science+Business Media B.V.
Deutscher G. (2005). The Unfolding of Language, London: William Heinemann
Gantt, E.E., Melling, B.S., and Reber, J.S. (2012). "Mechanisms or metaphors? The emptiness of evolutionary psychological explanations," Theory and Psychology, 22(6): 823-841.
Gibson, J.J. (2015). The Ecological Approach to Visual Perception: Classic Edition, New York, NY: Psychology Press. (original1986).
Giddens, A. (1984). The Constitution of Society: Outline of the Theory of Structuration, Cambridge: Polity Press.
Hayles,N.K (1999). How We Became Posthuman: Virtual bodies in Cybernetics, Literature, and Informatica, Chicago/London: University of Chicago Press.
Heylighen, F., and Joslyn, C. (2001). "Cybernetics and second-order cybernetics", in: R.A. Meyers (ed.), Encyclopedia of Physical Science & Technology (3rd ed.), New York: Academic Press.
Heylighen, F. Cilliers, P. and Gershenson, C., (2007). "Complexity and philosophy," in J. Bogg and R. Geyer (eds.), Complexity, Science and Society, Oxford: Radcliffe
Huang et al. (2020). "Drug development targetting SARS Cov 2," Acta Pharmacologica
Jablonka, E., and Lamb, C. (2005). "Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life," Cambridge, MA: The MIT Press.
Jörg, T. (2021). Generative Complexity in a Complex Generative World: A Generative Revolution in the Making, Cham, Switzerland: Springer.
Kim, D. (1999). Systems Thinking, Waltham, Mass: Pegasus Communications Inc.
Laland. K.N., Odling-Smee, J., and Feldman, M.J. (2000). "Niche construction, biological evolution, and cultural change," Behavioral and Brain Sciences, 23: 131–175.
Lettwin, J.Y., Maturana, H.R., Pitts, W., and McCulloch, W.S. (1959). "What the frog's eye tells the frog's brain," Proceedings of the Institute of Radio Engineers, 47: 1940-59. Reprinted in W. S. McCulloch, Embodiments of Mind. Cambridge, Mass.: MIT Press, 1965. (pp. 230-255)
Maturana, H.R. (1970). "Neurophysiology of cognition," in P. Garvin (ed.), Cognition: A Multiple View, New York: Spartan Books, pp 3-24.
Maturana, H.R. (1971). Biology of Cognition. BCL Report 9.0, Biological Computer Laboratory, Dept of Electrical Engineering. University of Illinois (mimeograph)
Maturana, H. (1978). "Biology of language: The epistemology of reality," in G. Miller and E. Lenneberg (eds.), Psychology and Biology of Language and Thought: Essays in Honor of Eric Lenneberg, New York: Academic Press, pp 27-64.
Maturana, H.R. (1988). "Reality: The search for objectivity or the quest for a compelling argument," Irish J. of Psychology (special issue on Radical Constructivism) 9(1): 25-82.
Maturana, H.R. (1990). "Science and daily life: The ontology of scientific explanations," in W. Krohn, and G. Kuppers (eds.), Self Organization: Portrait of a Scientific Revolution, Dordrecht: Kluwer Academic Publishers.
Maturana, H. (1997). Metadesign, Retrieved August 1, 2005, from http://www.inteco.cl
Maturana, H.R., and Frenk, S. (1963). "Directional movement and horizontal edge detectors in the pigeon retina," Science, 1963, 142: 977-9.
Maturana, H.R., and Frenk, S. (1965). "Synaptic connections of the centrifugal fibers in the pigeon retina," Science, 150(694): 359-61.
Maturana, H.R., and Guiloff, G.D. (1980). "The quest for the intelligence of intelligence," Journal of Social and Biological Structures, 3(2): 135-148.
Maturana. H., and Mpodozis, J. (2000). "The origin of species by means of natural drift," Rev. Chil. Hist. Nat., 73(2). Online Journal http://www.scielo.cl. Accessed 27th August 2009.
Maturana, H., Mpodozis, J., and Letelier, J.C. (1995). "The nervous systems as closed neuronal network; Behavioral and cognitive consequences," From Natural to Artificial Neural Computation: Lectures Notes in Computer Science, 930: 130-136.
Maturana, H., Mpodozis, J., and Letelier, J.C. (1995). "Brain, language and the origin of human mental functions," Biological Research, 28(1): 15-26.
Maturana, H., and Varela, F. (1980). Autopoiesis and Cognition, Reidal Publishing: Dordrecht, Holland
Maturana, H.R., and Varela, F.J. (1992). The Tree of Knowledge: The Biological Roots of Human Understanding, Boston: Shambhala.
Maturana, H.R., Verden-Zoller, G., and Bunnell, P. (2008). The Origin of Humanness in the Biology of Love., Exeter, UK: Imprint Academic.
Mingers, J. (1996). "A comparison of maturana's autopoietic social theory and Giddens' theory of structuration," Systems Research, 13(4): 469-482
Mitchell, M. (2009). Complexity: A Guided Tour, Oxford: Oxford University Press.
Montuori, A. (2008). "Editor’s foreword," in E. Morin (ed.), On complexity, Cresskill, N.J: Hampton Press.
Montuori, A. (2013). "Complexity and Transdisciplinarity: Reflections on theory and practice, world futures", The Journal of Global Education, 69: 4-6, 200-230.
Morin, E. (2008). On Complexity, Cresskill, N.J: Hampton Press.
Oyama, S. (2000). Evolution's Eye: A Systems View of the Biology-Culture Divide, Durham and London: Duke University Press.
Oyama, S., Griffiths, P.E., and Gray, R.D. (2001). Cycles of Contingency: Developmental Systems and Evolution, Cambridge, Mass: MIT.
Preiser, R., Cilliers, P. and Human, O. (2016). Deconstruction and complexity. in R. Preiser (ed.), Critical Complexity: Collected Essays, Berlin: Boston : De Gruyter
Pigliucci, M., and Müller, G.B. (2010). Evolution – The Extended Synthesis, MIT Press, Cambridge, Mass, USA
Prigogine, I. (1997). The End of Certainty: Time, Chaos, and the New Laws of Nature, New York: Free Press.
Prigogine, I., and Stengers, I. (1984). Order Out of Chaos: Man’s New Dialogue with Nature, London: Heinemann.
Senge, P.M. (2006). The Fifth Discipline: The Art & Practice of the Learning Organization, New York: Random House Business. (original 1990)
Shannon, C. (1948). "A mathematical theory of communication," in W. Weaver and C.E. Shannon (1963). The Mathematical Theory of Communication, Univ. of Illinois Press. Available at: ada.evergreen.edu/~arunc/texts/cybernetics/weaver.pdf. Retrieved October 2014
Thompson E. (2011). "Living ways of sense making," Philosophy Today, 55(Supplement): 114-123.
Turner, S. (2004). "Extended phenotypes and extended organisms," Biology and Philosophy, 19: 327–352, 2004.
Varela, F. (1979). Principles of Biological Autonomy, New York: Elsevier (North-Holland)
Varela, F. (1996a). "Neurophenomenology: A Methodological remedy to the hard problem," J.Consc.Studies 3: 330-350.
Varela, F. (1996b) "A science of consciousness as if experience mattered," in R. S. Hameroff, A. W. Kaszniak, and A. C. Scott (eds.), Toward a Science of Consciousness, MIT Press.
Varela, F.J., Thompson, E., and Rosch, E. (1991). The Embodied Mind: Cognitive Science and Human Experience, Cambridge, Mass: MIT Press.
Weaver, W., and Shannon, C.E., (1963). The Mathematical Theory of Communication, Univ. of Illinois Press. Available at: ada.evergreen.edu/~arunc/texts/cybernetics/weaver.pdf. Retrieved October 2014.
Weber, S., and Varela, F. (2002). "Life after Kant: Natural purposes and the autopoietic foundations of biological individuality," Phenomenology and the Cognitive Sciences, 1: 97–125, 2002.
Woermann, M. (2011). What's complexity theory? Features and implications, https://www.academia.edu/823563/What_is_complexity_theory. Retrieved 8.01.22.
Woermann, M. (2016). Bridging Complexity and Post-Structuralism: Insights and Implications, Springer International Publishing AG.
Wolstenholme, E.F. (1990) System Enquiry. A System Dynamics Approach, Chichester: John Wiley and Sons.